





TOP > Infinite dream > 岡部 幸司
破骨細胞の生理学、ダイナミックな生命現象を求めて!!
福岡歯科大学 細胞分子生物学講座 細胞生理学分野 教授 岡部 幸司
- 破骨細胞
- イオン動態
- パッチクランプ法

スロベニアで開催されたヨットのヨーロッパ選手権大会で、国際審判を担当しました。
この度は「骨研究のトップランナー」の紹介依頼を頂き、大変、恐縮しております。そもそも、私が骨研究のトップランナーかどうか、骨代謝領域の専門家かどうかも、背中がむず痒い気がします。とは言うものの、私が骨代謝研究を始めたきっかけや、これまで取り組みや研究に対する思い、趣味のヨットのことなどを徒然に述べてみます。これからの研究を目指す若い方達に少しでも参考になれば幸いです。
Gタンパクを介する受容体とイオンチャネル制御機構の研究
私は当初から骨代謝研究を始めていたわけではありません。私は1984年に九州大学の歯学部卒業後、九州大学医学部・薬理学教室の大学院に進み、平滑筋研究のトップランナーである栗山煕教授の下で血管平滑筋の研究に取り組みました。そこは医学部の大学院生30名を有する大きな研究室で、平滑筋の機能を筋収縮や電気生理学的手法、生化学的手法を取り入れて、あらゆる角度で研究が進められていました。私に与えられたテーマは、当時としては最新技術であったパッチクランプ法を用いて、酵素で単離した一個の動脈平滑筋細胞に直径1μmのガラス電極を吸着させて、イオン輸送を解析することでした。動脈平滑筋は弾性組織に埋まっていて単離するのが難しく、また、単離した動脈平滑筋細胞の表面も硬く歪で実験用チャンバーの底面にも接着し難く、パッチ電極を近づけるだけでごろごろと転がって逃げていくので、幾度も失敗を繰り返すうちに高度な技術が身に付きました。そして肺動脈平滑筋細胞からの世界で初めて平滑筋収縮に重要なCa2+チャネルの性質とCa拮抗薬の作用機序を解明することに成功し、これが私の学位論文となりました。大学院卒業後、テキサス州ヒューストンにあるBaylor College of Medicineの分子生理学研究室(Arthur Brown教授)に留学する機会を得ました。そこで心筋の自律神経調節に関わるムスカリン受容体—GTP結合タンパク質—K+チャネルのシグナル伝達の仕組みや3量体Gタンパクを構成する各サブユニットの機能解明に取り組みました。この研究室には世界中からありとあらゆる種類のGタンパクが集められており、この2年間の研究生活は、Nature、Cell 、Scienceと名だたるトップジャーナルへ向けた日々競争の大変厳しいものでした。アセチルコリンにより作動する単一K+チャネルを記録しながら各種Gタンパク質サブユニットとの機能的連動をミリ秒の時間オーダーでダイナミックに解析する「イオン電流を用いた生化学的実験」は、大変難易度の高いものでしたが、ここで大学院時代に動脈平滑筋で苦労して身につけた経験や技術が大いに役立ちました。今思えば、この2年間で10年分に相当する実験をこなした気がしますし、「どんな状況においても絶対やりきる精神力」が培われました。ここで私はアゴニストが結合していなくても受容体はGタンパクに対して「車のアイドリング」のように一定の情報を流しており、アゴニストが結合後の迅速な活性化に備えていることを発見しました。1990年に帰国後、山口大学医学部生理学教室の助手として妊娠や性ホルモンによる子宮平滑筋の機能変化とイオン輸送のメカニズムについて研究しました。その後、福岡歯科大学口腔生理学教室の副田教授からのお誘いで1993年から異動し、2002年よりは教室名を細胞生理学分野と改めると共に現職に就任しました。以上のように、私の研究ライフワークの前半部分は、受容体—Gタンパク—イオンチャネル制御と平滑筋や心筋の収縮機構に関するものなのです。

恩師の栗山煕先生が、私の教授就任祝賀会にご出席くださった時の写真。
破骨細胞との出会い
この様なこてこてのイオンチャネル屋であった私が骨代謝研究を始めたきっかけは、山口大学在職中に指導した整形外科の大学院生が「ラットの骨髄中には破骨細胞がいるんですよ、これ実験に使えませんか」と培養ディッシュにまいた破骨細胞を見せてくれたことです。初めて見た多核の破骨細胞の癒し系の姿に興味を引かれた私は、早速これまで腕を磨いてきた得意のパッチクランプ法を応用し、「それではイオン輸送機構を調べてみるか」と始めたのが骨代謝研究の始まりでした。パッチクランプ法の特徴は、個々の生きた破骨細胞の細胞膜に存在する一個のイオン輸送体の機能をダイナミックに解析できることで、顕微鏡下で見る実験用チャンバー内に目標の細胞が一個あれば実験できるのです。例えば、リン酸カルシウム上で骨吸収状態と非吸収状態の破骨細胞を選択することもできるので、パッチ電極を使った「破骨細胞の魚釣り」みたいな実験ですね。ご存じのように、破骨細胞は波状縁(ruffled border)からのプロトンポンプ(V-ATPase)を介するプロトン(H+)分泌とCl-輸送体を介するCl-分泌により、骨表面に強酸性の環境を形成し骨ミネラルを溶解します。まさに破骨細胞の役割はH+やCl-等のイオンを分泌することなのですが、これらを担うイオン輸送体のダイナミックな特性やこれらを調節する機構など生理的かつ本質的なことは意外と未だに分かっていないのです。
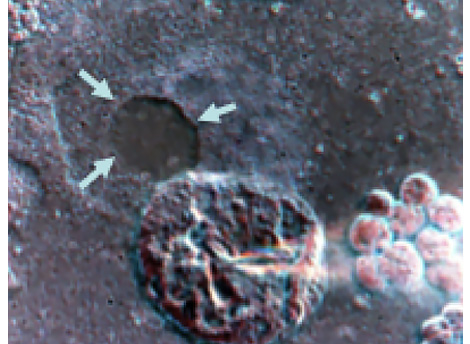
破骨細胞をパッチ電極で移動させると、細胞直下に吸収窩が認められますので、吸収状態の細胞だったといえますね。
破骨細胞のイオン動態と骨吸収調節に関する研究
福岡歯科大学に移ってからは、本格的に「破骨細胞のイオン輸送と骨吸収機構」について研究を始めました。有り難いことにスタッフの岡本富士雄先生、鍛治屋浩先生にも仲間に入ってもらい、イオン動態のイメージング解析や生化学的な手法も取り入れ、自見英治郎先生(現:九州歯科大学教授)がエール大学から准教授として赴任してからは、更に遺伝子改変動物等を用いた分子生物学的な方法論も充実し、少しずつ破骨細胞のイオン動態を多くの手法で多面的に研究する体制ができてきました。
これまで我々は骨吸収抑制ホルモンであるカルシトニンがPKA系を介して破骨細胞のCl-輸送やV-ATPaseやNa+/H+交換輸送を介するプロトン分泌を調節することや、酸分泌にはClC7型Cl-輸送体やK+-Cl-共輸送体を介するCl-分泌が重要であることを明らかにしてきました。また、欧米諸国で多い常染色体優性大理石骨病Ⅱ型(ADO Ⅱ)の患者で報告されたClC7 点変異が破骨細胞のCl-分泌を低下させることが病因であることも分かりました。さらに、ビスフォスフォネートの作用は一般的に時間をかけて骨吸収を抑制すると考えられていますが、パッチクランプ法を用いると、驚いたことに窒素含有型ビスフォスフォネートは、数分間の瞬時に破骨細胞のCl-分泌をメバロン酸代謝経路を介して抑制することが解りました。現在、このClC7とプロトンポンプ(V-ATPase)の両輸送体分子の間には、両者と複合体を形成し、それぞれの発現や機能調節を行う共通の分子が存在することが明らかとなっており、こういったカップリング分子を標的とした、より効率の良い創薬開発にも応用できるのではと考えています。
一方、骨吸収機構だけでなく破骨細胞の分化に関わるイオン機構にも取り組んでいます。破骨細胞の分化過程には、RANKL 刺激によって誘発されるCa2+オシレーションの発生、これに続くCa2+依存性脱リン酸化酵素(calcineurin)や転写因子NFATc1の活性化が重要であります。このCa2+オシレーションの形成を担う分子実体や形成機序について検討したところ、破骨細胞の分化初期過程において、RANKL 刺激により破骨前駆細胞にCa2+透過性の非選択性陽イオンチャネルであるTRPV2 の発現が上昇し,これが自発的で周期的なCa2+オシレーションを形成すると共に、その後NFATc1の活性化には細胞内Ca2+ストア機能(Stim1・Orai1分子)や PLC 系がカップリングすることが明らかとなりました。パッチクランプ法とCa2+イメージングを併用して、TRPV2を介するイオン動態と細胞内Ca2+と濃度変化の動的な連動を解析することができたのです。つまりTRPV2 は破骨細胞のCa2+オシレーションを構築する重要なキー分子であると言うことであります。

ラボのスタッフや大学院生、留学生と会食会。
研究への思いとこれからの展望
以上、破骨細胞のイオン動態に関する研究の一部をご紹介しましたが、私が基本的に知りたいのは、「破骨細胞に何ができるかではなく、破骨細胞が何をしているか」ですね。近年、研究手法は格段に進歩しましたが、遺伝子改変技術を含む多くの分子生物学的な尖端技術が駆使され、ややもすると、むりやり破骨細胞に何かさせているのではないかと感じる研究を見受けます。破骨細胞の本質的な生理学はまだまだ未解決なこが多いと思います、今後も生きている破骨細胞の骨吸収機構を分子レベルやダイナミックなイオン動態の面から、硬組織の病態解明に一石を投じる研究ができればと願っています。
一方、我々の研究室ではイオン動態の研究だけではなく、これまで破骨細胞分化に関わるPKAやNF-κBをはじめとした多くの細胞内シグナル伝達系による調節機構や癌骨転移機構、歯の萌出と歯根吸収調節機構をはじめ多くのテーマに取り組んでいます。昨年よりは福岡歯科大学出身である福島秀文先生が准教授として赴任し、ユビキチン化を介する破骨細胞分化や発がんの制御機構について新しい領域の研究を始めています。現在は、歯学部のテーマとして、象牙芽細胞やエナメル芽細胞に高発現するTRPチャネルに焦点をあて、ミネラルイオンの動態や遺伝子改変動物を用いて歯牙の形成や石灰化機構についても研究を行っており、この成果に大いに期待をしています。
私は浪人して大学へ進学し、その後も多くの節目には失敗ばかりでした。学生時代はヨットばかりやっていて、どうにか試験はぎりぎりでクリアしていたのですが、生理学の試験だけは再試験を受けました。その時、必至で勉強したのがきっかけで、柄にもなく生理学の研究をしてみたいなと思ったのです、私が大学院に進学する時、同級生には「え、本気か?」と言われましたね。それが、今は学生に生理学の神髄を教授しているのですから、巡り合わせとは皮肉なものです。私が留学する時に尊敬する先生から「恐れず、いっぱい失敗をしてきなさい。それが必ず将来につながるから。」と送り出されましたが、その通りだったと思います。今では、研究が思うようにいかなくても、それは失敗ではなく計画を軌道修正するのだと前向きに考えるようにしています。これまで一生懸命に取り組んできまたが、就職活動せずとも、大学院卒後の留学、帰国後の就職、福岡歯科大学への異動の折々には、何れもご指導頂いた多くの先生方のお導きで職を得てどうにか現在に至っております。これからも人と人の出会いや繋がり、感謝の気持ちや初心を忘れずに納得のいく研究を目指したいと思います。

大阪の居酒屋で、ビリケンさんを囲んで。右から、自見先生、私、福島先生。
生涯スポーツとしてのヨット競技
九州大学の学生時代、全学のヨット部に所属し、大学生活のうち4年間は、年間135日位ですかね、土日と学校が休みの時は全てヨット部の合宿、お陰で年中日焼して真っ黒で、白衣とのコントラストが鮮明で病院実習が恥ずかしかったですね。実は、九大ヨット部は日本で最初にできた大学ヨット部で、創部86周年を迎える大変伝統のあるヨットクラブなのです。学生時代の戦績は全日本インカレ9位でした。その後、ヨットは生涯スポーツとして現在も続けています。幾度か日本代表として世界選手権にも出場し競技選手としてだけではなく、国体、インカレ、インターハイや世界選手権など国内外の大会で審判員や大学ヨット部の監督として、また日本の審判委員を育成する講習会の指導者として今でもヨットに情熱を注いでいますし、東京オリンピックに向けての選手育成と開催支援ができればと願っています。ただ、ヨットというアウトドアスポーツは遊んだのがごまかせないので、学生からは「こげん色の黒か教授はおらん」とか、さらに、教授会では「どげんしたら、そこまで日焼けできるとですか?」と相変わらず呆れられております。結局は、ヨットが好きで、海が好き、私にとって、大学雑務や研究から開放され気分転換できる魔法のスイッチでもあります。今後は体力の衰えに抵抗しながら、年齢に応じたヨットライフ(特に日焼け対策は入念に)をおくっていきたいと念じております。

本年度の全日本歯学生体育大会(オールデンタル)で、指導する本学のヨット部が団体戦と個人戦の完全優勝を果たしました。