





TOP > Infinite dream > 小竹 茂
破骨細胞研究取り組んできた24年間
東京女子医大膠原病リウマチ痛風センター 小竹 茂
- 破骨細胞
- IL-17
- TCTA
- VDAC

私たちの研究メンバー。
2012年11月8日、東京女子医大膠原病リウマチ痛風センターの研究室にて。
1 はじめに
『記憶してください。私はこんな風にして生きてきたのです。』
今年、岩波書店から出版後100年目にあたる夏目漱石の『こころ』のなかで「先生」が「私」に手紙の中で語る部分です。このホームページの私の研究歴がどの程度、皆様の参考になるかわかりません。しかし、兎に角以下に述べたように私は研究を進めてきました。実験と違って、人生にはコントロールがなく、しかも完全な『一回性』です。ですから、正しかったか、そうでなかったかは、誰にもわかりません。常に、その時に、ベストと考える方向へ突き進んでいくだけです。骨代謝学に興味がありこれから研究をはじめる方、特に臨床をやりながら研究を行おうとしている方、には多少なりとも参考になることを期待して、以下に私の研究歴を述べてみます。「記憶して」いただかなくても結構ですので、最後まで、お読みいただければ幸いです。
2 留学前
私の学生時代は、多田富雄、利根川進、本庶祐の3氏が同時に朝日賞を受賞するというような、免疫学全盛の時代でした。私は、免疫学に興味をもつ同学年の何人かと勉強会などを行っていました。1987年に東京医科歯科大学医学部を卒業し、第一内科に入局しました。3年間の内科学全般の臨床研修を終了し、臨床の専門はリウマチ学にきめ医科歯科にもどり宮坂信之先生の指導のもとリウマチ学の臨床を始めました。当時は卒後4年目で専門を決めていたのです。1990年の秋頃からは医科歯科の檜垣恵先生、女子医大の佐藤和人先生の指導のもとin vitroの研究を始めました。檜垣先生には実験ノートの書き方から指導を受けました。『その日の天候も含め詳細に記載すること』といわれたことは今でもよく覚えています。初めに行った実験は関節リウマチ(rheumatoid arthritis, RA)患者の脛骨の骨髄血をメチルセルロースという粘度の高い培地で培養するコロニー・アッセイでした。この実験は医科歯科の血液内科の奈良信雄先生に指導いただきました。さらに、奈良先生には、実験結果が非常に重要でそこから考察することの重要性を教えていただきました。その実験の結果CFU-GMがRA骨髄には増加しているということを明らかにしました。これが私の初めての英文原著論文となり1992年、J Rheumatolに掲載されました。
その頃、私は外来にてRA患者で関節が溶けてしまうような関節変形が非常に高度な、いわゆるムチランスタイプの患者を診察しました。当時のRAの病因・病態研究はHLA研究、B細胞やT細胞などの免疫系の細胞、滑膜細胞、あるいはIL-1などの炎症性サイトカインなどの研究が主流でした。しかし、私は、関節の変形をまずは、なんとかすべきであり、それには破骨細胞の研究が非常に重要だと考えるようになりました。当時、RAの病態において破骨細胞に関してはほとんど研究されていませんでした。もともと学生時代から免疫学に興味があり、特に単球/マクロファージが好きな細胞でした。そこで、日本内科学会、日本リウマチ学会はもちろんですが、日本免疫学会、日本臨床免疫学会、日本骨代謝学会などにも所属し、免疫学的な視点から破骨細胞の研究を始めようと考えました。1993年にRAの滑膜組織中にauthenticな破骨細胞を見いだしアメリカリウマチ学会(ACR)において発表しましたが、破骨細胞関連の発表は私の1演題のみでmiscellenous(種々雑多)の領域に入れられてしまっていました。それほど、当時は、破骨細胞研究はRA研究の中では、『異端』だったのです。
当時、破骨細胞の前駆細胞は単球/マクロファージであり、そのまた前駆細胞はCFU-MまたはCFU-GMということはわかっていました。このような流れからも、破骨細胞に興味をもち破骨細胞の研究を始めました。破骨細胞は多核の細胞で骨に張り付いて骨組織を削っていくという非常に特徴のある細胞で、独特の魅力があります。私はその魅力に取り憑かれてしまいました。破骨細胞研究者は、自嘲ぎみに(あるいは自慢げに?)自らのことを『破骨細胞オタク』といいますが、わたしも自覚することなく、いつしか、その『破骨細胞オタク』になってしまっていたようです。
私は、1991年頃から、様々な関連論文を読みながら、東京女子医大の膠原病リウマチ痛風センターの研究室でヒトの末梢血から単核球を分離し、様々なサイトカインを添加し培養して多核のTRAP陽性細胞の形成に成功しました。当時、破骨細胞ができたとよろこんだのですが、後からTRAP染色はin vivoでは特異性が高いがin vitroではマクロファージ系の細胞もTRAP陽性になることを知りました(Modderman WE et al. Tartrate-resistant acid phosphatase is not an exclusive marker for mouse osteoclasts in cell culture. Bone. 1991)。ですから、この多核のTRAP陽性細胞はマクロファージのpolykarionの可能性が大きかったのだと思います(ただし、骨吸収活性の確認はしていませんでした)。しかし、今にして思えば複数のサイトカインの相乗作用を期待するのは、あながち的外れでもなかったのではと思っています。
当時のin vitroでの破骨細胞形成は昭和大学の高橋直之先生らのマウス由来細胞を用いた共存培養系が確立されていました。このままでは私の破骨細胞研究は先に進まないと考え、佐藤和人先生に昭和大学の口腔病理学教室の山口朗先生をご紹介いただき、破骨細胞と骨芽細胞の二重染色法などを教えていただきました。さらに山口先生から、当時は破骨細胞研究の『メッカ』であった昭和大学の須田立雄先生の口腔生化学教室をご紹介いただきました。それ以降、須田先生の研究室(以下、『須田ラボ』)に通うようになり、高橋直之先生、宇田川信之先生に破骨細胞の培養方法などの指導をうけました。先ほど説明したヒトTRAP陽性細胞を宇田川先生に見せたとき『これは、誰が、どうやって培養したのですか?』と、質問されたことを今でも鮮明に覚えています。
須田ラボは非常に活気があり当時、宮浦先生、東大整形の田中栄先生の後にいらしていた仲村一郎先生、高見先生、自見先生、茶木先生など、あるいは企業の研究者など、まさに『梁山泊』でした。名前を「さん」で呼びあうのも、臨床の医局での生活での感覚からはとても新鮮でした。私は、この須田ラボで、基礎の教室の厳しさと魅力を実感することができました。一方、女子医大の膠原病リウマチ痛風センターの研究室ではRAの滑膜細胞の培養や免疫染色を行っていました。RAの関節液中には可溶性のIL-6受容体が増えていることを初めてみいだし、関節液中のIL-6と可溶性のIL-6受容体により破骨細胞形成が促進することも明らかにしました(JBMR 1996)。
3 留学
1994年4月からは米国NIHのDr. R. L. Wilder(図2左)のラボに留学しました。図2の中央はDr. Wilderの奥様、右は私です。ラットの関節炎モデルの遺伝子解析が中心テーマのラボでした。私はそのラットの研究も行いながら、発症早期関節炎の滑膜組織のサイトカインprofileを、nested RT-PCRの方法を確立するところからはじめました(J Immunol Methods, 1996;Proc Assoc Am Physicians, 1997)。

(図2)NIHの私のボスのDr. R. L. Wilder(左)、Dr. Wilderの奥様(中央)、そして私(右)。1995年。
さらに、当時Pensilvania大学のDr. Schumacher H. R. Jr.(図3)が、サバティカルでNIHにて反応性関節炎の一つであるクラミジア関連関節炎の研究を行っていました。Dr. Schumacher H. R. Jr.は、クラミジア関連関節炎研究の泰斗であり、当時アメリカリウマチ学会の重鎮でした。この先生からクラミジア関連関節炎研究の指導もうけました(Infect Immun, 1999)。余談ですが、我が国にはこの疾患の臨床医が非常に少なく、この論文のおかげで、日本在住の外国人が私の外来に時々受診してくれています。また、私が脊椎関節炎も臨床で専門にするようになったのは、この研究がきっかけでした。

(図3)Dr. Schumacher H. R. Jr.
この留学期間中は破骨細胞研究からは離れてしまいました。しかしNIH での研究のシステムがよくわかりました。ラボのトップはアメリカ人で実際実験しているのは、ほとんどが中国、東欧、南米、日本からの留学生です。日本人は一定期間後帰国するので余裕があるのですが、その他の国からの留学生はアメリカに残ることを目指しているので留学生同士の競争は熾烈でした。私の留学生活は、研究以外で、あまり悩むこともなく、家族で旅行もよくできましたし、世界各地からきた研究者仲間との交流も楽しく、さらに、日本人研究者との知り合いも多数できました。ノーベル賞受賞者を多数排出している世界最大級の研究所で約3年間研究に従事できたことは、とても有意義でした。
4 留学後
1)IL-17
1996年12月に帰国しました。再び須田ラボに通うようになり、当時、須田ラボと雪印研究所との共同で、OPG, RANKL(当時の須田ラボではODFと命名)の発見のまっただ中でした。私はIL-17の研究を始めました。IL-17は破骨細胞の分化を促進させることを見いだしました(図4)。さらに当時RAはTh1疾患とされていましたが、滑膜組織にはT細胞が多数存在しているのにIFNγが検出されにくいことが謎とされていました。私はT細胞由来のサイトカインであるIL-17を通常のELISAで容易に検出でき、しかも濃度が上昇しており、RAの病態にIL-17が重要な役割を果たしていることも明らかとしました(図4)(J Clin Invest 1999)。このように、IL-17が破骨細胞の分化に関与していること、さらに関節リウマチの病態にも関与していることを私たちが世界に先駆けて報告しました。2005年には新たなヘルパーT細胞であるTh17が発見されました。この論文は非常に多く引用され続けており、これまで1158回(2014年8月14日現在) 引用されています。IL-17研究は免疫学では一つの大きな研究領域として急速な勢いで発展中です。生物学的製剤として抗IL-17抗体の治験も複数進行中です。ただし、この治験はTNFα阻害療法に効果がなかった患者を対象としていることが私は問題だと考えています。私は、IL-17は関節リウマチの早期患者の病態に非常に重要であると考えています。現在、厚生労働省の研究班で発症早期関節炎患者におけるTh17の研究を推進しています。さらに私たちは、IL-17は骨芽細胞が存在しない条件でもヒト破骨細胞の分化を促進すること(Yago et al. J Cell Biochem, 2009.)、Th17の分化に関与しているIL-23がヒト破骨細胞の分化を促進すること(Yago et al. Arthritis Res Ther, 2007)なども明らかにしています。
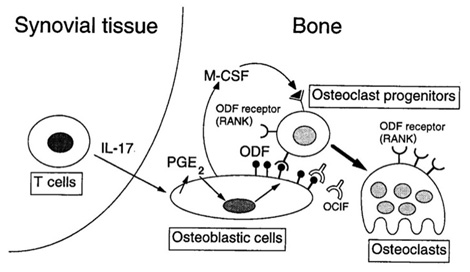
(図4)滑膜のメモリーT細胞はIL-17を産生し、関節液のIL-17は通常のELISAで検出できる。IL-17はマウス骨芽細胞のRANKL産生を促進し、マウス単球からマウス破骨細胞を形成する。
Kotake et al. J Clin Invest 1999
IL-17研究の次に、私は、T細胞がRANKLを発現していることから、T細胞が直接的に単球/マクロファージを破骨細胞に分化させ、RAの病態に関与している、という仮説をたてました。この仮説のもと検討した結果、この仮説を支持する結果を得ることができました(Arthritis Rheum 2001)。なお、マウスの細胞で他の複数の研究グループも同じ頃に同様の結果を報告しましたが、ヒトの細胞で示したのは私が初めてでした。
2)T cell leukemia translocation-associated gene (TCTA)
1996年の帰国直後から始めたもう一つの研究は、関節リウマチ滑膜から、ヒト破骨細胞の分化を抑制あるいは促進させる新規分子を発見するプロジェクトでした。滑膜組織から蛋白を精製し、その破骨細胞分化への活性を測定し、この作業を繰り返し最終的には、TCTA蛋白が単球/マクロファージ、破骨細胞の細胞膜に発現し、分化を促進させていることを明らかにしました。(Bone 2009)(図5)興味深いことに,このTCTA蛋白由来ペプチドはヒト肺小細胞ガンの分化を抑制することも明らかにしました(J Cancer Ther, 2013)。
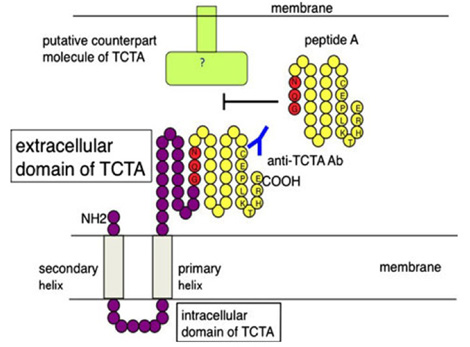
(図5)T-cell leukemia translocation-associated gene (TCTA)蛋白:破骨細胞分化に重要な分子。TCTA由来のペプチドAがTCTA蛋白の機能を阻害する。
Kotake et al. Bone 2009
3)Voltage-dependent anion channel (VDAC, porin)
留学後に始めた、もう一つの研究は、ヒト破骨細胞から膜蛋白を抽出しそれに対するモノクローナル抗体を作成し、ヒト破骨細胞の膜の新規蛋白を発見するというプロジェクトです。質量分析法なども駆使し、最終的にCl-のイオンチャンネルであるVDACがヒト破骨細胞膜に発現していることを見いだしました(Cell Biol Int. 2013)(図6)。VDACは通常はミトコンドリアの外膜に発現しています。破骨細胞はプロトンを産生し骨を吸収する細胞ですが、電気的にバランスをとるために陰イオンであるCl-チャンネルも非常に重要で、すでに複数のCl-チャンネルが報告されていました。私は、VDACは新たなヒト破骨細胞のCl-チャンネルであるということを明らかにしました。抗VDAC抗体はヒト破骨細胞の分化を抑制するということを、科学技術機構(JST)の支援のもと、国際特許に出願中です。
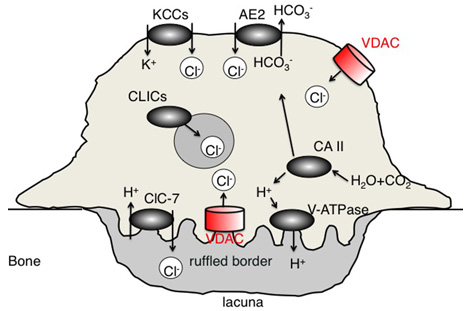
(図6)破骨細胞の新規の膜Cl-イオンチャンネルであるvoltage-dependent anion channel (VDAC, porin)を同定。
Kotake et al. Cell Biol Int 2013
4)既存薬の破骨細胞に対する効果
私たちは、これまでテプレノン(セルベックス®)(Nanke et al. Calcif Tissue Int, 2005)、タクロリムス(プログラフ®)(Yago et al. Cytokine. 2012)、レバミピド(ムコスタ®)(南家ほか、第32回日本骨代謝学会学術集会2014、第35回日本炎症・再生医学会2014)などがヒト破骨細胞の分化を抑制することを報告してきました。
5)『ヒト破骨細胞学 Human Osteoclastology』
マウスではIFNγは破骨細胞の分化を強力に抑制することが報告されていましたが、私たちはヒトIFNγ産生T細胞はRANKLをも発現し、ヒト破骨細胞の分化を促進することを見いだしました(Eur J Immunol 2005)。また、齧歯類(げっしるい)の門歯はヒトとは異なり伸び続けます。この門歯は物をかじることで次第に削れてゆき、長さを保っています。この『齧(げっ)』は『齧る(かじる)』から由来しているわけです。さらに、前述のように、私の最初の研究は血液学的手法のコロニー・アッセイでした。CFU-Mは破骨細胞の前駆細胞であり、マウスM−CSFはCFU-Mのコロニーを形成します。しかし、ヒトM−CSFはCFU-Mのコロニーは形成せず分化させるのみです。この破骨細胞の分化に非常に重要な点においてマウスとヒトでは全く異なります。また前述のTCTA研究の時にもヒトとマウスの違いが明らかになりました(Bone 2009)。以上のような事実などから私は2008年に5年間の私立大学戦略的研究基盤支援事業に採択された時に『ヒト破骨細胞学 Human Osteoclastology』という用語を考案しました。この用語はその後、アメリカリウマチ学会の公式雑誌Arthritis RheumのEditorial (Arthritis Rheum, 2009)を私が執筆するときに紹介し、とても好評でした。その後、引用が年々増加してきています。なお、私は平成22年度に日本骨代謝学会の学術賞(内科系)をいただきましたが、その時の依頼英文総説(J Bone Miner Metab. 2012)もご参照いただけますと幸いです。もちろんマウスの実験は重要であり、私たちもマウスを使用し実験を行ってきています。しかし、ヒトとマウスの違いを常に考え研究を行うことを強調していきたいと考えています。
5 私の趣味
学生時代には大学のオーケストラでバイオリンを弾いていました。1987年に研修医になってからは多忙で、バイオリンは中断していました。しかし、2000年から再開し、医局の忘年会では私の研究グループのメンバーでバイオリン、ピアノ、フルート、パーカッションなどでのアンサンブルを余興として『古典音楽同好会』と自称し、クラシックの名曲を6年間連続で演奏していました。また、私は、毎年バイオリン教室の発表会で演奏していました。好きな作曲家はJ.S.バッハとブラームスです。これまで、ブラームスのバイオリンソナタ第3番にも挑戦してきました(図7)。2006年には西東京市の市民オーケストラである西東京交響楽団を中心に形成されたオーケストラによるモーツァルト記念演奏会にての『レクイエム』の演奏に1stバイオリンで参加しました。図8は学生の時から愛用しているバイオリンで、ストラディバリウスと同型に約100年前に作製されたバイオリンです。弓は4年前に新しくしました。

(右:図7)バイオリン演奏中の私です。
(左:図8)愛用のバイオリンと弓。バイオリンはストラディバリウスと同型に約100前に作製されたバイオリンです。
6 おわりに
研究は結果が出るか出ないかは始めて見ないとわかりません。どのような研究テーマに取り組むかは、私の経験からは、自分に興味があることが第一であり、他の人がやっていない領域に取り組むのがいいと考えています。好きなことであれば時間をかけることは苦ではなく、むしろ楽しく、結果がでればそれはとても良いことですが、でなくてもそれはそれであきらめもつくと思います。現在、流行の領域に参入するのは、いわゆる『バンドワゴン現象』といって避けた方がよいと思います。