





TOP > Infinite dream > 今泉 和則
やってみないとわからない-渡り歩いてたどり着いた先が骨研究
広島大学大学院医歯薬保健学研究院分子細胞情報学 今泉 和則
- 小胞体
- 転写因子
- 骨芽細胞
- 軟骨細胞

今から25年程前、大阪大学医学部解剖学第二講座(遠山正彌教授)でアルツハイマー病研究を始めたのが実質的な研究人生のスタートでした。アルツハイマー病は脳内で神経細胞死が生じ認知機能が低下する病気です。当時、家族性アルツハイマー病の原因遺伝子としてプレセニリン1が発見され、その機能と病態形成の分子機構に注目が集まっていました。優秀な研究者の多くは、βアミロイド産生におけるプレセニリンが果たす役割について研究を進めていましたが、私は世界のライバルたちと同じ土俵で戦っても勝ち目はないと考え、プレセニリン遺伝子の変異がなぜ神経細胞死を起こすのかという一点縛りで実験を行うことにしました。方法は非常に原始的です。遺伝子変異をもつプレセニリンを構成的に発現する細胞と野生型の細胞を準備し、ありとあらゆるストレス(酸化ストレス、低酸素ストレス、低グルコース、無血清、カルシウム濃度変化、各種の薬物刺激等)を加えた際のストレス感受性を調べるというものです。
当時、私は大学でのポストを持たない、企業から派遣された一介の研究員でしたが、知り合いや研究仲間を無理やり募って抄読会を企画、開催していました。その中でたまたま出会ったのが、「小胞体ストレス」という概念を酵母の系で提唱したCellの論文でした。何やらこのストレスが強く生じると酵母では細胞死を起こすらしい、その分子機構は?哺乳類でも起こるのだろうか?など、まだ数少ない小胞体ストレスの論文を読むにつれ興味が高まっていきました。一方で、運よくプレセニリンの良い抗体が作成でき、細胞を染めてみると、何とその多くのシグナルは小胞体に集積していたのです。そこで細胞に小胞体ストレスを負荷したところ、変異プレセニリンを発現する細胞が劇的に死んでいきました。他の様々なストレスでは野生型との差が見出せなかった実験で、初めてポジティブな結果が出て興奮したものです。その後は、細胞レベルではありましたが、分子機構をほぼ解明しNature Cell Biologyに掲載していただきました。
私はアルツハイマー病の病態発症機構を解明したくてこのような研究を行ってきましたが、小胞体ストレスという概念に触れてからは、むしろこちらの研究に傾倒するようになりました。というのは、シグナル経路を含め余りにも未知な点が多かったからです。特にシグナル発信分子である小胞体ストレスセンサーの可能性にも魅力を感じていました。酵母ではIRE1という小胞体ストレスセンサーが見つかっていたものの、哺乳細胞ではひとつも見つかっておらず、哺乳細胞における新たなセンサーを求めて研究を開始しました。結果的には先を越されて、3つのメジャーなストレスセンサーがDavid Ron、Randal J. Kaufman、そして日本の森和俊先生により発見されました。このときは非常に落胆しました。それでもまだセンサーは有るという信念(ただの思い入れだけ)から、新たなセンサーの同定にチャレンジし続けました。なぜ、そのように錯覚のような信念があったかというと、私は小胞体ストレスの実験を繰り返す中、3つのセンサーだけでは説明ができない非常に奇妙な現象を何度も見ていたからです。そのひとつは、神経細胞とグリア細胞の共培養系に小胞体ストレスを負荷すると、神経細胞は速やかに細胞死を起こすのですが、グリア細胞はそう簡単には死にません。
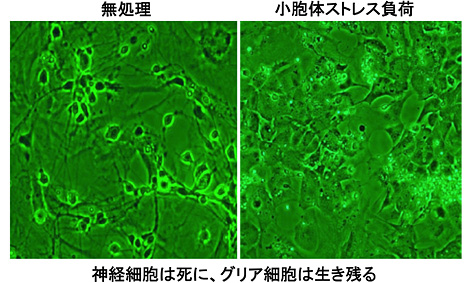
神経細胞は死に、グリア細胞は生き残る
おそらくグリア細胞だけ小胞体ストレス下でも生き残る何かが備わっている、すなわち新たなセンサーあるいは神経細胞にはないシグナル経路がグリア細胞に備わっているからだと考えるようになりました。また、内分泌系の細胞では大量のタンパク質が合成・分泌されており、そのような細胞では常時小胞体ストレス状態になっています。しかし非分泌系の細胞では通常小胞体ストレスはかかっていません。3つの主要な小胞体ストレスセンサーは基本的にはどの細胞でも同じ程度発現しています。分泌系の細胞が常に小胞体ストレスに曝されているにも関わらず、細胞死を起こさず生存していることも、3つのストレスセンサーだけでは説明がつかないと考えていました。細胞・組織特異的な新たなセンサーあるいはシグナル分子を探して、毎日探索作業に明け暮れました。
色々行った中で唯一上手くいったのは、データベースサーチでした。当時見つかっていた小胞体ストレスセンサーおよびその下流シグナル分子のデータベース解析を徹底的に行ったところ、センサーのひとつであるATF6という分子に機能ドメインの配置が比較的類似した分子を見つけることができたのです。膜貫通型でしかも転写因子。小胞体に局在してもおかしくない構造をもつ5つの分子(OASIS、BBF2H7、Luman、CREBH、AIbZIP)です。すぐさまタッグ付きの発現ベクターを作成し細胞内での局在を調べたところ、5つとも見事に小胞体に局在していることがわかり、小胞体ストレスセンサーの可能性が高いことが確認できました。5つの分子の中から、まずは神経系に発現するOASISとBBF2H7にフォーカスして研究を進めました。いずれも小胞体ストレスに応答して膜内切断を受け転写因子として働くことを証明し、Nature Cell BiologyやMolecular Cellular Biologyに報告しました。その後はin vivoでの機能解析へと展開させていったのです。
OASIS欠損マウスを作成したところ、脳には一見何も変化が見られなかったことから大きな失望感に苛まれました。野生型マウスと見た目が違うのは、手足が若干短く細いかなという程度でした。“手足じゃなぁ・・・”というのがその時の素直な感想でした。学生時代、製薬企業在籍時代とも病理学が専門でしたので、昔取った杵柄ではないですが、取り敢えず全身の組織を病理組織学的に隈なく観察することにしました。結局何も変化を見出せなかったのですが、最後に脱灰した骨組織の顕微鏡観察をすると、骨梁がやせ細り骨密度も著しく低下、皮質骨も薄くなっていることに初めて気付いたのです。[図3]
私は骨の解析など全く行ったことがなかったものですから、この先どう進めればよいか戸惑い解析を止めてしまおうかとも考えました。師匠の遠山先生に相談したところ、「骨やったら米田に聞いてみい!」とアドバイスをいただきました。失礼にも「誰ですか?その人?」とお尋ねしたところ、骨代謝学会の理事長で、阪大歯学部教授の米田俊之先生だと教えていだきました。ちなみに遠山先生とは同級生で、卓球部でもご一緒だったらしいです。その後は米田先生や当時米田研の准教授でいらした西村理行先生から懇切丁寧にご指導をいただき、瞬く間に骨形態計測、骨芽細胞の培養系を用いたin vitro実験、電子顕微鏡解析等を完了させ、OASISが骨形成に必須であることを証明することができました。
OASISと並行して進めていたもうひとつのセンサーBBF2H7のノックアウトマウスも神経系には何も変化がなく骨格系に異常が見られました。こちらの場合は、軟骨細胞の小胞体機能が障害されたことで軟骨マトリックスの分泌が阻害され軟骨形成不全を起こします。[図4]
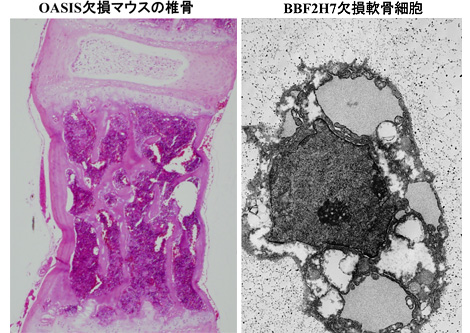
[上:図3]OASIS欠損マウスの椎骨
[下:図4]BBF2H7欠損軟骨細胞
つまりBBF2H7は軟骨マトリックスの分泌と軟骨形成に必須の分子であったのです。やはり軟骨の解析も自分たちだけではできなかったので、米田先生、西村先生から一般的な軟骨解析ツールをご提供いただき、また培養法等たくさんの技術指導も仰ぎました。両先生なくして私の骨・軟骨研究は在り得なかったと今でも思っています。そうこうしているうちに、次々と骨領域にお知り合いでき、骨を研究する環境が整ってきました。今では骨・軟骨解析でできないことはないと思えるほど、様々なテクニックを研究室のメンバーが習得してくれています。そして、今走らせている研究テーマの半分以上が骨領域をベースとしたものになっています。
以上、長々と書いてきましたが、研究領域を定めないで、必要に応じて新たな分野にチャレンジしてきた私のやり方が果たして良かったのかどうかはわかりません。ボトムアップ式に特定の領域の研究を積み上げて行く姿勢もその領域研究を発展させるには極めて重要だと思われます。ひとつ言えることは、興味がある現象や思い入れのある分子を追究したいと感じた時には、“案ずるより産むが易し“で、領域の壁を越えて未知に挑もうとする姿勢が大切ではないかということです。私は研究室のメンバーにいつも「まずはやってみないとわからんやろ」と激励しています。チャレンジングな研究に挑む若い研究者がどんどん増えていくことを願って止みません。

研究室のメンバー
なお、小胞体ストレスと骨・軟骨形成に関しては総説をいくつか執筆しています(生化学84:18-29, 2012、実験医学31:849-855, 2013等)ので、そちらのほうも是非ご覧下さい。
最後に私の趣味は、高校時代からの仲間と良く行くゴルフです。ヘタの横好きで、人に見せられるものではありませんが、研究と同じように、スイングの軌道やインパクト時のヘッドの入射角など微に入り細を穿って追究しております。スコアは一向に良くなりませんが・・・・

趣味のゴルフ
研究所住所
〒734-8553
広島市南区霞1-2-3
広島大学大学院医歯薬保健学研究院分子細胞情報学
Email: imaizumi@hiroshima-u.ac.jp