





TOP > Brave heart > 高橋 直之
私の破骨細胞研究
松本歯科大学総合歯科医学研究所 高橋 直之

はじめに
私は昭和大学と松本歯科大学において破骨細胞の研究を行い、その成果を日本骨代謝学会で報告してきた。骨代謝に関する様々な知識を日本骨代謝学会の学術講演やシンポジウムそして出版物等で学んだ。私はこの学会に研究者として育てられたと実感している。日本骨代謝学会のホームページへの寄稿を依頼されたときには大変に名誉なことであると嬉しく思った。2020年3月に私は松本歯科大学を退職したが、その際「破骨細胞に魅了されて」という退職記念誌を作製した。本稿では、その記念誌をもとに私が破骨細胞に出会った経緯とどのように破骨細胞を研究してきたかを記し、私の研究者としての足跡をお示ししたい。
1.岩手大学時代
私は1976年岩手大学農学部農芸化学科を卒業後、大学院修士課程に進学した。農芸化学科は土壌学、微生物学、栄養化学、生物化学、農産製造学などの研究室があるが、私は発展性を秘めていると魅力を感じ、生物化学教室を選んだ。当時、勝又悌三教授と江尻慎一郎助教授が研究室を運営していた。勝又先生は、植物の生殖を生化学的に解明しようとしていた。一方、江尻先生は蛋白質合成機構の解明に取り組まれていた。私は勝又先生の研究班に入り、アカマツ花粉の花粉管の伸長に関わる環状ヌクレオチドの役割の研究に従事した。修士2年目の夏に、私は生化学関係の雑誌に須田立雄先生の教室の助手の募集を見た。須田先生は昭和大学歯学部生化学教室の初代教授として、東京医科歯科大学から1977年に赴任された。須田先生の書かれた総説を読みビタミンDがホルモンに変わるということを知り興味を持った。研究が面白そうだと思えてきたことから、もう少し研究に従事したいと思い、須田先生に願書を送った。幸運にも須田先生の教室の助手に採用された。須田先生は「高橋君は歯学部出身でもないから、まず数年間一緒にやってみましょう。そのあと、高橋君も進路を考えてください。」と話された。私も当時はそう長いこと歯学部に籍を置くことはないだろうと考えていた。

農芸化学科生物化学教室の方々
昭和大学への就職が決まった1977年の秋に須田先生は、私にMundy先生の論文を読むようにと指示された。その論文は、多発性骨髄腫が破骨細胞活性化因子を分泌しているという内容だった。ビタミンDに関連する論文ではなかったために、不思議に思ったことを覚えている。須田先生はビタミンDの骨吸収促進作用に興味を持っていたことを後に知った。私は破骨細胞がどのような細胞かも知らずにこの論文を読んだが、後にMundy先生の教室に留学し、生涯のテーマとなる破骨細胞研究に入った。不思議な巡り合わせがあるものだと思う。
2. 昭和大学における研究生活
◆ 最初に行った研究
ビタミンDは小腸のCa吸収を促進するホルモンである。須田先生の教室で私が最初に行った研究は、鳥類のCa代謝の解明を目指すものであった。産卵期の鳥類は、輸卵管の子宮部で炭酸Caよりなる卵殻を形成する。そのため、そのため膨大部では大量のCaが輸送される。「このCa輸送にビタミンDが関わるかを明らかにすること」が与えられたテーマであった。助教授の堀内 登先生の指導を受け、ウズラを用いて実験を開始した。産卵期のウズラは、毎日産卵する。私はまず、ウズラの輸卵管子宮部にビタミンD受容体(VDR)が存在することを明らかにした。産卵期のウズラをビタミンD欠乏食で飼育すると、数日内に卵殻の石灰化が低下した軟卵を産む。輸卵管子宮部にVDRの存在を確認していたので、子宮部でのCa輸送が障害されたものと予想した。産卵期の鳥類は、長管骨の骨髄の中に骨髄骨という独特な骨組織を形成し、卵殻形成に必要なCaを蓄える。卵殻形成期(夜間)に破骨細胞が骨髄骨を吸収してCaを供給する。産卵後に破骨細胞は活動を停止し、骨芽細胞が骨髄骨の再形成を開始する。産卵期の鳥類は毎日これを繰り返す。ビタミンD欠乏食で飼育したウズラを調べると、子宮部でのCa輸送の障害というより、むしろ骨髄骨の石灰化不全が軟卵を産む原因であることが判明した。ビタミンD欠乏により、小腸のCa吸収が低下し、その結果骨髄骨は未石灰化骨になっていた。破骨細胞は未石灰化骨を吸収することができないために、軟卵を産むという結論になった。この研究において、昭和大学歯学部病理学教室の山口 朗先生から、骨組織の形態学を教えていただいた。研究が面白く思えるようになったのはこの頃である。
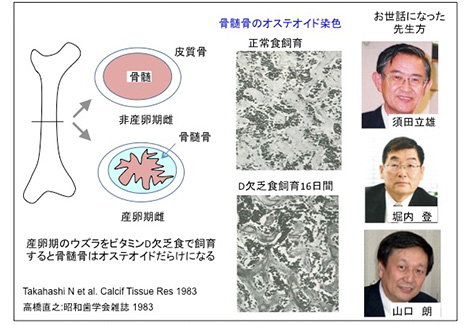
学位論文: 産卵期のウズラのCa代謝に関する研究
◆ Mundy先生とRoodman先生
1984年に、「産卵期のウズラのCa代謝に関する研究」で東京医科歯科大学より歯学博士の学位が授与された。学位を取得したことで、須田先生から留学許可が下りた。須田先生の勧めもあり、岩手大学時代に読んだ破骨細胞論文の著者、テキサス大学San Antonio校のGreg Mundy先生の教室に留学した。Mundy先生はCa代謝の内分泌学を専門とする研究者で、米田俊之先生(大阪大学歯学部)がすでに留学されていた。当時Mundy先生の教室は、悪性腫瘍が分泌する高Ca血症惹起因子を同定しようと教室一丸で取り組んでいた。この因子は、後にJack Martin先生らによりPTHrPと同定された。私は、高Ca血症惹起因子研究のプロジェクトに入りたかった。しかしMundy先生は、この因子が破骨細胞の形成を促進することから、私を同僚のDavid Roodman先生に紹介し、破骨細胞の研究をするように薦めた。Roodman先生は血液腫瘍学の研究者で、骨髄細胞培養系を用いて、破骨細胞形成を試みていた。一方、須田教室の阿部悦子先生は、活性型ビタミンDが肺胞マクロファージから多核細胞の形成を誘導することを見出していた。須田先生たちはこの多核細胞が破骨細胞ではないかと考えていた。そのため、私は須田教室と競合するような破骨細胞研究を行いたくなかった。しかし、Roodman先生が親身に私たち家族を世話してくださったこともあり、留学期間中は破骨細胞研究に専念しようと決めた。ヒト骨髄細胞を長期間培養し、破骨細胞形成を観察した。活性型ビタミンDや副甲状腺ホルモンを添加すると破骨細胞様の多核細胞が誘導される。その過程を解析して、いくつかの論文を執筆した。

Mundy先生とRoodman 先生
◆ 久米川先生と破骨細胞の映像
1986年7月に留学を終え、8月に昭和大学に復帰した。その直前1986年6月に米国骨代謝学会学術集会がアナハイムのディズニーランドホテルで開かれた。私の発表が口演に選ばれたこともあり、記憶に残る学会であったが、それにもまして忘れられないことがある。明海大学の久米川正好先生は、この学会に破骨細胞の映像フィルムを持参していた。学会会場となったホテルの一室を借り、久米川先生が持参された16 mmフィルムの上映会が行われた。10人ほどが重なり合ってその映像を見たが、そこにはマウスの骨髄細胞由来の多核細胞が骨を吸収している像が映し出されていた。私は、骨髄胞培養系で形成される多核細胞が骨を吸収する破骨細胞であるか不安に思っていたが、この映像をみて、帰国後はマウスの骨髄細胞培養系を用い破骨細胞形成に取り組もうと決意した。
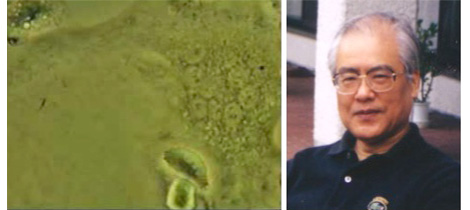
マウス骨髄細胞培養系に出現した多核細胞と久米川好正先生
◆ 共存培養系の確立
私は帰国後すぐに、マウス骨髄細胞の培養実験に取り掛かった。須田先生は、肺胞マクロファージ由来の多核細胞は破骨細胞ではないと結論していたために、私の破骨細胞研究を支援してくださった。マウスの骨髄細胞の培養系において、活性型ビタミンDにより誘導される多核細胞は、骨吸収能をもつ破骨細胞であった。また、破骨細胞は必ず骨芽細胞様の間質細胞に接して形成された。そこで、私たちは破骨細胞の分化には骨芽細胞が必要ではないかと考えるようになった。一方、Gideon Rodan先生とJack Martin先生は、PTHやPGE2などの骨吸収促進因子は骨芽細胞に作用して破骨細胞を骨組織に誘導するという仮説を1981年に報告した。また1983年に、新潟大学の小澤英浩先生の教室の大学院生だった江尻貞一先生は胎仔ラットの頭頂骨を形態学的に観察し、破骨細胞の分化に骨芽細胞が関与する可能性を示した。そこで私たちは、骨芽細胞と脾細胞(破骨細胞の前駆細胞は含まれるが骨芽細胞様間質細胞は含まれない)の共存培養系を確立した。活性型ビタミンDを添加すると、多数の破骨細胞の形成が認められた。この共存培養系では、様々な骨吸収促進因子が破骨細胞形成を促進したことから、破骨細胞の分化に骨芽細胞が関わることが広く認められるようになった。共存培養系の確立は、防衛医科大学校の大学院生赤津拓彦さんと、須田教室の大学院生宇田川信之さんの協力なしにはなされなかった。昭和大学において、以下のような破骨細胞研究が行われた。
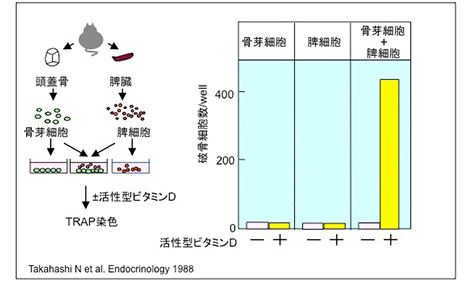
骨芽細胞と脾細胞の共存培養系での破骨細胞形成
◆ M-CSFと肺胞マクロファージ
1990年にop/opマウスの原因遺伝子(M-CSF)が判明し、M-CSFは破骨細胞分化に必須なサイトカインであることが明らかにされた。一方宇田川さんは、骨芽細胞様の骨髄間質細胞株ST2細胞とMC3T3-G2/PA6細胞が造血細胞との共存培養系で破骨細胞形成を支持することを見出した。さらに、単球やマクロファージもST2細胞との共存培養系で破骨細胞に分化できることを報告した。この実験結果は、大きな反響を呼んだ。破骨細胞研究の第一人者であるTim Chambers 先生はマクロファージから破骨細胞を作る実験が再現できず、彼の教室の大学院生を須田教室に送りたいとの手紙も届いた。最終的には、Chambers先生らもマクロファージから破骨細胞を形成させることに成功し、破骨細胞前駆細胞は単球・マクロファージ系であることが認められるようになった。東京大学医学部整形外科学教室の大学院生だった田中 栄さんは、M-CSFの作用を詳細に解析し、M-CSFは破骨細胞の前駆細胞の増殖のみならず分化にも必須であることを証明した。
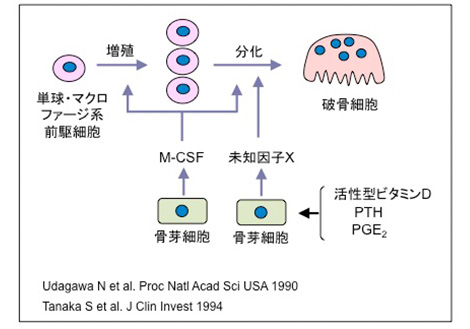
破骨細胞の分化におけるM-CSFの役割
◆ IL-6と可溶型IL-6受容体
IL-6やIL-11はシグナル伝達因子gp130を使って標的細胞内にシグナルを伝達する。共存培養系においてIL-11やOncostatin Mは強力に破骨細胞形成を促進したが、IL-6は破骨細胞形成を誘導しなかった。IL-6受容体は、単球など破骨細胞前駆細胞は発現するが、骨芽細胞は発現していない。その頃、大阪大学の岸本忠三先生たちは、可溶性IL-6受容体は、細胞がIL-6受容体を発現していなくてもgp130にIL-6シグナルを伝達できることを発見した。そこで私たちは、岸本先生から可溶性IL-6受容体を供与していただき、共存培養系に添加した。可溶性IL-6受容体を入れると、IL-6は濃度依存的に破骨細胞形成を促進した。この研究は、中外製薬の田村達也さんと宇田川さんによって行われた。宇田川さんはさらにヒトIL-6受容体を発現する遺伝子改変マウスを用い、骨吸収促進因子は骨芽細胞に作用して破骨細胞を誘導することを明らかにした。
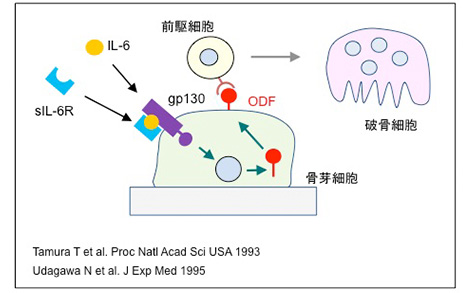
IL-6 は可溶型IL-6受容体を利用してODFの発現を誘導する
◆ 破骨細胞は骨の硬さを認識する
破骨細胞は骨面に波状縁と明帯を構築して、骨面に酸と蛋白質分解酵素を分泌する。これは「破骨細胞の極性化」と呼ばれる。破骨細胞が骨の何を認識して極性化するかは不明であった。多くの研究者は、破骨細胞は骨基質蛋白質を認識して波状縁を形成すると考えてきた。東京大学医学部整形外科学教室の大学院生だった仲村一郎さんは、破骨細胞の極性化機構に興味を持っていた。仲村さんは様々な仮説を立てて実験を行い、最終的に「破骨細胞は接着した骨の堅さを認識して極性化する」ことを証明した。破骨細胞は骨の硬さを認識し明帯と波状縁を形成すると結論した。さらに、この極性化にはp130Casと呼ばれるキナーゼが関わることを示した。後に、東京大学医学部整形外科学教室出身の澤田泰宏先生は、p130Casが硬さを認識するforce-sensing kinaseであることを証明した。
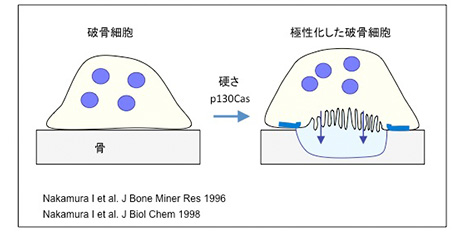
破骨細胞は硬さを認識し極性化する
◆ 破骨細胞活性化因子とIL-1
九州大学歯学部の大学院生だった自見英治郎さんは、破骨細胞におけるNF-κBの役割を研究するために昭和大学に来た。自見さんは、破骨細胞はIL-1受容体を発現しており、IL-1は破骨細胞を活性化することを見出した。富士裾野で開かれた「Bone Cell Biologyの会」で、IL-1作用についてChambers先生と議論していた自見さんを思い出す。さらに自見さんは、骨芽細胞は破骨細胞を活性化する因子を発現していることを見出した。破骨細胞活性化因子と名付けたが、最終的には、この因子の実態はRANKLであることが後に判明した。
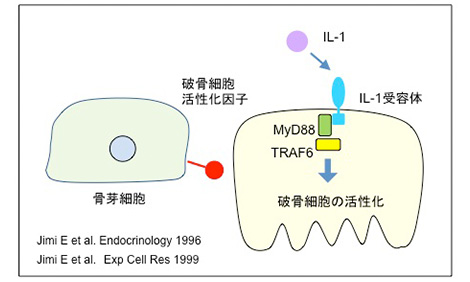
骨芽細胞とIL-1は破骨細胞を活性化する
◆ 破骨細胞分化因子ODF仮説とRANKLの発見
私たちは1992年に、破骨細胞分化因子(osteoclast differentiation factor, ODF)に関する一つの仮説を発表した。その仮説では、①骨吸収促進因子は骨芽細胞に作用する、②骨芽細胞はODFを細胞膜上に発現する、③破骨細胞前駆細胞は細胞間接触を介してODFを認識し、M-CSFの存在下で破骨細胞に分化する、というものであった。
1997年、Amgenの研究者は新規の骨吸収抑制因子としてOPG (osteoprotegerin)を発見した。一方、雪印乳業の研究者も同時期に破骨細胞形成を抑制する新規因子OCIF (osteoclastogenesis inhibitory factor)を発見した。OPGとOCIFは同一因子で、TNF受容体ファミリーに属するが、膜貫通ドメインを持たない分泌型蛋白質だった。私たちは、OPG/OCIF はODFのデコイ受容体であると想定した。雪印乳業との共同研究より、ODFは活性型ビタミンDで処理されたST2細胞からOCIF結合因子としてクローニングされた。一方Amgenの研究者もOPG結合因子をクローニングし、その因子をOPG ligand (OPGL)と命名した。私たちがクローニングしたODFは、既報のTRANCEあるいはRANKLと同一因子であった。
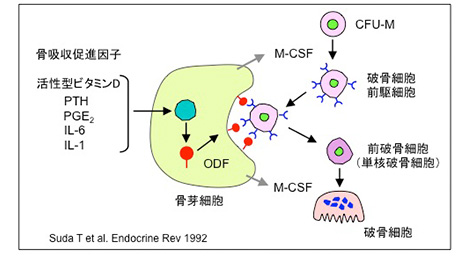
破骨細胞形成におけるODF仮説
破骨細胞の形成を誘導するリガンドにはODF, OPGL, TRANCE そしてRANKLという4つの名前が付けられていた。一方ODFのデコイ受容体には、OPGとOCIFという2つの名前が付けられていた。受容体は、RANKであることが確認された。2000年の米国骨代謝学会で、学術名命名会議が開かれた。私たちは、ODFがふさわしいと主張したが、リガンドをRANKL、デコイ受容体をOPG、そして受容体をRANKと呼ぶように推奨することが決まった。後にRANKLは乳腺幹細胞、胸腺髄質上皮細胞、そして小腸パイエル板M細胞の分化に必須なサイトカインでもあることが示された。ODFではなく、RANKLという名前でよかったと私は考えている。
◆ 雪印乳業の研究者と高柳 広先生
東尾侃二先生が率いた雪印乳業の研究者たちは、精悍な研究者集団であった。1997年の春に、雪印乳業の研究者たち(東尾侃二先生、保田尚孝先生、津田英資先生)との話し合いが、昭和大学で行われた。須田先生、宇田川さん、そして私がその話し合い臨んだ。そこでODFのクローニングに成功したことが告げられた。脾細胞のみの培養系でリコンビナントODFにより誘導された多核細胞は、共存培養系で観察された破骨細胞と全く同様であった。後日、保田尚孝先生から雪印乳業の小さな研究集団がAmgen社よりも研究と特許で先行できたのは、「焦点を絞って研究したからである」と伺った。研究において、焦点を絞ることの重要性を学んだ。
私たちと同様にRANKLの発見を知った多くの研究者は、破骨細胞研究はこれで終わったと大きな喪失感を感じた。一方、東京大学医学部整形外科学教室出身の高柳 広先生は、この発見を見て、「破骨細胞研究はここから始まる」と胸を高まらせという。その後の高柳先生のご活躍を見ると、「ここから始まる!」という心構えが常に大切であることを学んだ。

昭和大学の仲間たち
3. 松本歯科大学における研究
2001年松本歯科大学の小澤英浩先生から、「松本歯科大学に新しい大学院を作りましょう」というお誘いを受け、宇田川信之先生と共に松本歯科大学に赴任した。小澤先生は、新潟大学の口腔解剖学教室の教授を務められたあと、松本歯科大学口腔解剖学の教授として2000年に赴任された。のちに松本歯科大学の学長と初代研究科長を務められた。松本歯科大学は宇田川先生の母校でもあることから、極度に緊張することもなく松本歯科大学に赴任できた。松本歯科大学創立者である矢ヶ﨑 康先生は「立派な大学院を作りましょう」と歓迎してくださり、研究者を集めることや総歯研の研究室の設計などを私たちに任せてくれた。松本歯科大学の教員と職員の皆さんも私たちを温かく迎えてくれた。松本歯科大学の皆様のご支援には、心より感謝する。
小澤英浩先生は2000年に、宇田川先生と私がその翌年に松本歯科大学に赴任した。これが契機となり、小澤英浩先生、成人病診療研究所の白木正孝先生、そして信州大学医学部整形外科学教室教授の高岡邦夫先生が代表世話人となって、「松本ボーンフォーラム」が結成され、宇田川先生と私がその事務局を担当した。「松本ボーンフォーラム」に多くの硬組織研究者をお迎えすることができ、私たちは多くの課題を深く学ぶことができた。2002年に高岡先生は、大阪市立大学に赴任され、そのあと信州大学医学部整形外科学教室教授になられた加藤博之先生が代表世話人を引き継がれた。この会を支援してくださった帝人ファーマ株式会社に深謝する。骨粗鬆症治療においても各職種の専門性を生かしたチーム医療が行われるようになってきた。その流れを受けて、2019年からは「松本ボーンフォーラム」は「信州骨代謝多職種セミナー」として引き継がれている。
私と宇田川先生は破骨細胞に魅了されているために、松本歯科大学で行われた研究も骨吸収に関するものが中心であった。松本歯科大学で行われた研究を以下に紹介する。

松本歯科大学の仲間たち
● 骨代謝共役因子(カップリングファクター)
破骨細胞が吸収した骨は骨芽細胞が埋め戻す。そこには、骨代謝共役(カップリング)機構が働くと考えられているが、その実態は不明である。中村美どり先生は最初にOPG欠損マウスを用いて骨代謝共役機構を解析した。OPG欠損マウスは、骨吸収と骨形成がともに高度に亢進しているマウスであることを明らかにした。次にOPG欠損マウスにビスフォスフォネートを投与し、骨吸収を抑制すると骨形成も見事に抑制された。私はこの結果を見て、精緻な骨代謝共役機構が存在することを確信した。
スクレロスチンは骨細胞が分泌しWntシグナルを阻止することで骨形成を抑制する骨形成抑制因子である。小出雅則先生は、破骨細胞は骨細胞のスクレロスチンの発現を低下させる因子を分泌していることを見出した。さらに破骨細胞の分泌するその因子はLIFであることを示した。この結果をみて、私は骨代謝共役機構の一端が明らかにされたことを実感した。一方山下照仁先生は、スクレロスチンの遺伝子座にGFP遺伝子を入れたSOST-GFPマウスの作製に成功した。このマウスを用いることで、骨代謝共役機構の解明がさらに進むことが期待される。
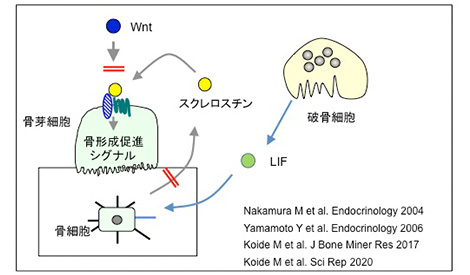
破骨細胞は骨細胞を介して骨形成を促進する
● 歯周病の機序とその治療
私たちは、歯周炎における歯槽骨吸収に関する研究も開始した。歯周炎を惹起する細菌の成分であるリポ多糖(LPS)の破骨細胞形成誘導機構を解析した。愛知学院大学の大学院生だった佐藤信行さんは、LPSは骨芽細胞のMyD88経路を介してRANKLを誘導することを示した。また、大学院生の堀部寛治さんは、CRAMP (ヒトの場合LL-37) と呼ばれる抗菌ペプチドはLPSの作用を阻害し、破骨細胞形成を抑制することを見出した。小出雅則先生はOPG欠損マウスは歯周病モデルであることを示し、ビスフォスフォネートや抗RANKL抗体は歯周病治療薬になり得ることを報告した。また、大学院生の尾﨑友輝さんは、W9と名付けられたペプチドは、RANKLに結合して破骨細胞分化を抑制するとともに骨芽細胞の分化を促進すること見出した。このように、歯周炎における歯槽骨吸収機構とその治療に関する研究が松本歯科大学で行えたことをうれしく思っている。
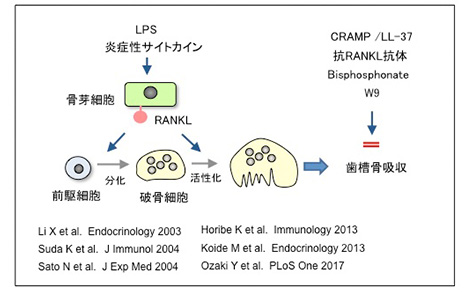
歯周炎における歯槽骨吸収誘導機序とその防御
● 破骨細胞前駆細胞の同定
骨芽細胞がRANKLとM-CSFを発現するため、破骨細胞は骨組織にのみ出現すると考えられてきた。しかしそうではない。愛知学院大学より派遣された大学院生山本洋平さんは、RANKL欠損マウスに大量のRANKLを投与して、破骨細胞の形成部位を観察した。破骨細胞は正確に骨組織にのみ形成された。またop/opマウスにM-CSFを投与しても、破骨細胞は正確に骨組織にのみ誘導される。以上の知見はRANKLとM-CSFの発現部位が破骨細胞の形成部位ではないことを示す。そこで、破骨細胞が骨組織に形成される機序を解析した。溝口利英先生は、破骨細胞の前駆細胞は細胞周期が停止した細胞であることを突き止め、静止期破骨細胞前駆細胞qOP (quiescent osteoclast precursors)と名付けた。愛知学院大学の大学院生武藤昭紀さんは、qOPは破骨細胞への分化がすでに運命決定された前駆細胞であること、その前駆は造血組織で形成されることを示した。また荒井 敦さんはこの運命決定にc-Fosがかかわることを証明した。IL-34はM-CSF 受容体に結合し、M-CSFと同様に作用する因子として発見された。中道裕子先生は、op/opマウスの場合、qOPは脾臓組織に存在すること、そしてIL-34は脾臓におけるqOPの維持に必須な因子であることを示した。さらに、op/opマウスにM-CSFを投与すると脾臓のqOPは血流を介して骨組織に運ばれそこで破骨細胞に分化することを明らかにした。現在私は、骨組織に運ばれたqOPは骨芽細胞にさらに教育され反応性破骨細胞前駆細胞rOP(responding osteoclast precursor)に分化すると考えている。
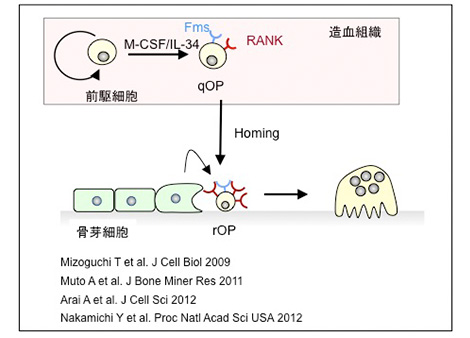
破骨細胞前駆細胞の動態
● 骨吸収とWntシグナル
Wntは骨代謝を調節する重要な因子で、そのシグナル経路はβ-カテニンを介する古典経路とそれを介さない非古典経路に分けられる。小林泰浩先生は、骨吸収を調節するWntシグナルを解析し、破骨細胞の中ではWnt非古典経路が重要な枠割を演じていることを明らかにした。慈恵会医科大学整形外科学教室の大学院生前田和洋さんは、Wnt5a-Ror2シグナルは破骨細胞前駆細胞のRANK発現を誘導し破骨細胞への分化を促進することを突き止めた。上原俊介先生は、Wnt5a-Ror2シグナルはRho→Pkn3→cSrcというシグナルの流れで破骨細胞を活性化すること、そしてPkn3阻害剤は骨粗鬆症モデルで骨量減少を防止できることを示した。一方、Wnt16はWnt5aと協調して骨吸収を調節することも示された。信州大学医学部整形外科学教室の大学院生岡本正則さんは、Wnt5a欠損の骨芽細胞は、Lrp 5/6の発現が低下するため、Wnt-β-カテニンシグナル伝達が損なわれていることを示した。このようにWntシグナルが骨吸収を調節していることを世界に先駆けて示すことができた。
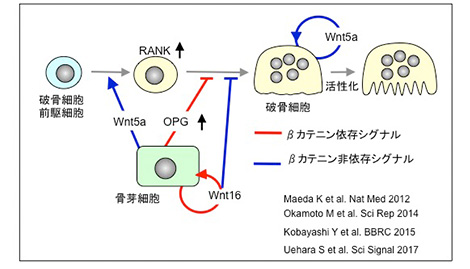
骨吸収を調節するWntシグナル
● ビタミンDの骨吸収作用
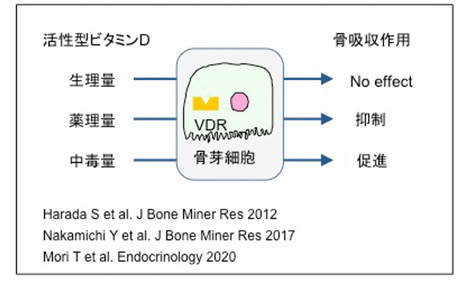
私は、須田先生のもとで研究を開始したこともあり、ビタミンDの骨作用にとりわけ興味を持ってきた。活性型ビタミンDは破骨細胞形成を促進する骨吸収を促進する。その一方で、活性型ビタミンD製剤[カルシトリオール、アルファカルシドール、エルデカルシトール]は骨吸収を抑制し、骨量を増加させる。このビタミンDの異なる骨作用の機序の解明を試みた。中外製薬の原田 卓さんと溝口利英先生は、マウスへのエルデカルシトールの薬理量投与は、骨芽細胞のRANKLの発現を低下させて骨吸収を抑制することを示した。一方中道裕子先生は、骨芽細胞特異的VDR欠損マウス(Ob-VDR-cKOマウス)を作出して、エルデカルシトール投与による骨吸収の抑制は、Ob-VDR-cKOマウスでは起こらないことを示した。同様の結果は、活性型ビタミンDの薬理量投与でも認められた。大学院生の森 智紀さんは、骨吸収を誘導するような中毒量の活性型ビタミンDを正常マウスとOb-VDR-cKOマウスに投与した。正常マウスへの活性型ビタミンDの中毒量の投与は骨吸収を伴うビタミンD中毒症を発症させた。しかし、ビタミンD中毒症はOb-VDR-cKOマウスでは起こらなかった。一方、生理量の活性型ビタミンDは骨吸収に影響を与えなかった。このように、薬理量の活性型ビタミンD投与は骨吸収を抑制し、中毒量の活性型ビタミンD投与は骨吸収を促進するということが示された。不思議なことに二つの相反する作用は骨芽細胞のVDRが担っていた。骨芽細胞のVDRの重要性を示すことができうれしく思っている。また、ビタミンD研究の面白さを再確認した。
活性型ビタミンDの骨吸収作用
おわりに
私はテキサス大学で破骨細胞に出会い、その魅力に取りつかれて、松本歯科大学に赴任してからも破骨細胞を研究してきた。研究が面白く、また論文を執筆することが楽しくてこれまでやってきた。私は、研究者は自らが知りたいことや解決したいことをテーマにするのがよいと思う。関連論文を読むことは楽しく、新しいテクニックの真剣に学びたいと思うだろう。若い人にはぜひ研究を楽しみ、研究生活を充実したものにしてほしいと期待する。何年か過ぎた後にお会いした時、「こんなに面白いことになっているんですよ!」と、笑顔で話しかけてほしいと願っている。
現在サイエンスから離れた生活となったが、サイエンスロスをそれほど感じていない。松本市を離れる前年の夏、孫娘と木工教室に通い、本棚、椅子、子犬の花置きを作った。昭和大学時代から時間を見つけては本棚や机などを作っていたが、松本歯科大学に赴任してからは、木工から距離を置いてきた。木工教室で木肌のぬくもりと木工の楽しさを思い出した。退職後はウッドデッキなどの大きな作品も作りたいと最後の一年間は工具を集めるなど準備をしてきた。現在それを実践している。木工作業も研究に通じるものがあり、創意と工夫が常に求められる。木工作品を作り上げる過程は、研究や論文執筆と似ており、完成形が見えてくるにつれてうれしさが増す。当分は楽しめそうである。今後は「先生」ではなく「棟梁」と呼ばれることを目指したいと考えている。
多くの方々のご指導とご支援を得て、私はこれまで研究者としてやってくることができました。皆様に心より感謝します。有り難うございました。

木工に魅了されて