





TOP > Brave heart > 須田 立雄
私と骨代謝研究の出逢い-『ビタミンDと骨』の研究の50年を振り返って
埼玉医科大学 客員教授 須田 立雄
私が大学(東京医科歯科大学・歯学部)を卒業したのは1960年のことである。それからもう50年以上が経過した。この度、日本骨代謝学会が我が国の骨代謝研究の黎明期を支えてきた先人達の業績を次世代に引き継ぐためのホームページを企画し、その一人として私にも寄稿するよう依頼があった。私は自分の能力も顧みずに、ただサイエンスが好きというだけの理由で「ビタミンDと骨」の研究に半世紀の時間を費やし、生涯を終わろうとしている一研究者である。 今回のお誘いは人生を締めくくる時期に来ているわが身の回顧のために良い機会であると前向きに捉え、原稿執筆をお引き受けすることにした。
私がこの50年間で行ってきた研究の中で、今でもそれなりに世間に評価されている仕事は、
[1] ビタミンD代謝の解明と活性型ビタミンD3の構造決定、
[2] Alfacalcidol [1α(OH)D3] の創薬研究、
[3] 活性型ビタミンDの細胞分化誘導作用の研究、
[4] 破骨細胞分化因子(ODF/RANKL)の研究
の4点であろうと思う。[1]は3年間の修業時代の研究で、私はただDeLuca先生の研究チームの一員として従事した仕事に過ぎない。[2]は米国から帰国して東京医科歯科大学助教授時代(6年間)の研究、[3]と[4]は独立して昭和大学歯学部で自分のラボで行った22年間の研究である。これらの研究を行う上で、日本骨代謝学会と米国骨代謝学会(ASBMR)は文字通り私の最も大切な研究発表の場となった。私のささやかな経験がこれから研究をスタートしようとする若い研究者に何らかの参考になれば望外の幸せである。
1.修業時代
私の生家は江戸時代末期から続く臨床歯科医の家系で、長男として生まれた私は当然家業を継ぐと父は期待していたし、私も漠然とそのように考えていた。しかし、大学は歯学部のない東京大学に進学し、2年後、父の要請もあり東京医科歯科大学・歯学部へ転学した。私の同級生にはその後「骨の形態学」の研究で有名になる小澤英浩君(新潟大学名誉教授)がいた。当時、東京医科歯科大学の基礎教育は歯学部と医学部が一緒になって行われることが多く、私は臨床研究よりも基礎研究に興味を持ち、大学院では「生化学」を専攻した。
「ビタミンD」との出逢いは最初の恩師となった荒谷真平先生(歯学部生化学教授)によって齎された。ある日、荒谷先生は日本橋の丸善でW.F. and M.W. Neuman 夫妻の著したThe Chemical Dynamics of Bone Mineral (University of Chicago Press, 1958) という単行本も見つけてきた。これは面白そうな本なので皆で翻訳しないか、ということになり、1960 年にその日本語版が「骨の生化学」というタイトルで医歯薬出版から刊行された[図1]。

(図1)私に「ビタミンDと骨の研究」を始めるきっかけを与えてくれた一冊の本―Neuman先生夫妻の「The Chemical Dynamics of Bone Mineral」(1958)
この本の最終章では骨の形成と吸収に関するNeuman夫妻の仮説が述べられており、この章を読んで私は「骨とビタミンD」の研究に挑戦してみたいと考えるようになった。 当時、すでに明らかにされていたビタミンDの標的臓器は小腸、腎臓、骨の3つで、ビタミンDはこれらの標的臓器でCa輸送を促進し、副甲状腺ホルモン(PTH)とともに血清Caの恒常性維持の主役を演ずる重要な因子と考えられていた。骨組織については、ビタミンDはPTHと共に骨から血清へCaを輸送する作用(骨吸収)を促進することはわかっていたが、血清中のCaが骨に沈着するプロセス(骨の石灰化)を直接促進するかどうかは不明であった[図2]。
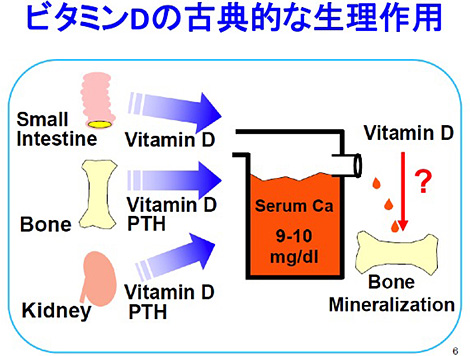
(図2)ビタミンDの古典的な生理作用
私はビタミンD研究の当時の専門家を調べて、Wisconsin 大学 (米) のH.F. DeLuca 先生の名前を知った。Wisconsin 大学には、医学の歴史上、ビタミンD研究の著名な科学者が多数輩出していて、水溶性ビタミン(water-soluble A)と脂溶性ビタミン(fat-soluble B) の発見者であるElmer McCollum 先生 (1879-1967) と、クル病の予防にB型紫外線 (UV-B) が重要であることを見出したHarry Steenbock先生(1886-1967)がおられたことも知った[図3]。

(図3)ビタミンD研究でWisconsin大学を有名にした3人の科学者
私はDeLuca先生に自分の留学希望を伝え、1年間待たされたが1968年7月からDeLuca先生のラボに留学できることになった。私より前にDeLuca 先生のラボに在籍していた日本人研究者の中には、ビタミンD3の最初の代謝産物(ピークIV, 後に25(OH)D3と同定される)を発見した森井浩世先生(大阪市立大学名誉教授)がおられたことを知り、留学前に森井先生ご夫妻にお逢いしていろいろなことを教えていただいた。
Wisconsin 大学に留学した2年7か月は私にとって生涯忘れることのできないサイエンス研究の修業の場となった。当時、DeLuca 先生のラボではビタミンDの代謝研究が精力的に行われていた。ライバルはケンブリッジ大学(英)のEgon Kodicek先生であった。 栄養素として摂取されたビタミンD2(3) と、UV-B によって皮膚の表皮細胞で産生されるビタミンD3は、当時は仮想的な代謝産物と考えられていた「活性型ビタミンD」に代謝されて、初めて生理作用を発現されるのではないかと考えられていた。
私がDeLuca先生のラボで研究をスタートした年(1968年)、ビタミンD3はまず肝臓に運ばれ、そこで25位が水酸化されて25(OH)D3に代謝されることが明らかにされた。この代謝産物は森井先生が発見されたピークIVであった。当初、DeLuca先生の研究室では25(OH)D3こそが活性型ビタミンDの本体であると思っていたが、英国のKodicek先生がピークPと名付けたシリカゲルのクロマトグラフィーでピークIVよりも移動度が遅い(水に溶けやすい)代謝産物が活性型ビタミンDであると報告された(1970年)。Kodicek先生のピークPは私たちがピークIVよりも遅れて溶出されるので、ピークVと呼んでいた代謝産物に一致した。当時、私たちはピークVが小腸で産生されると考えていたので、大学の近くにあったニワトリの加工工場から3万羽のニワトリの小腸を入手し、それを研究所の地下室にあったパイロット・プラントで処理して、最終的に2 μg の純化したピークVを得た[図4]。これが、現在活性型ビタミンD3の本体と考えられている1α,25(OH)2D3である(1971年)。

(図4)30,000羽のニワトリの小腸から活性型ビタミンD を単離・同定する大作戦
(a) ニワトリの加工工場、(b) 30,000羽のニワトリから集めた500kgの小腸サンプルの処理、
(c) Flash evaporator による小腸サンプルのEthanol抽出液の濃縮、(d) Sephadex LH20 column chromatography による最終精製
ほぼ同じころ、活性型ビタミンDは腎臓で産生されることがKodicek先生達により明らかにされ、今日理解されているビタミンDの代謝経路の全容が明らかになったのである。
すなわち、食物から摂取されたビタミンD2(3),あるいは皮膚の表皮細胞で作られたビタミンD3は血液中に存在するvitamin D-binding protein (DBP) に結合してまず肝臓に運ばれる。肝臓では肝細胞のミクロソームに存在するP450酵素(CYP2R1)によって25位が水酸化され、25(OH)D3 が生成される。25(OH)D3はDBPに結合して腎臓に運ばれ、DBPの受容体 (LRP2、Megalin) が豊富に存在する腎臓の近位尿細管細胞でエンドサイトーシスによって細胞内に取り込まれる。DBPから解離した25(OH)D3はミトコンドリアに取り込まれ、1α-水酸化酵素(CYP27B1)によって1α位が水酸化され1α,25(OH)2D3となるか、24-水酸化酵素 (CYP24A1) によって24位が水酸化され24,25(OH)2D3となる。このうち1α位の水酸化が活性化反応、24位の水酸化は不活性化反応である。1α,25(OH)2D3が血流を介して小腸、腎臓、骨などのビタミンDの標的臓器に運ばれると、核内受容体 (VDR) に結合し、ビタミンDの標的遺伝子のvitamin D response element (VDRE) を介して転写を促進する。そのようにして転写誘導されたタンパク質がビタミンDの不活性化酵素(CYP24A1)や小腸のCa輸送タンパクcalbindin であることも明らかになった[図5]。
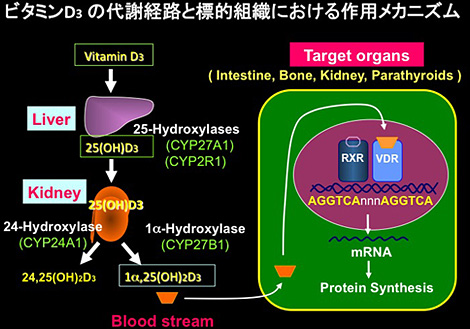
(図5)明らかにされたビタミンD3の代謝経路と活性型ビタミンDの標的臓器における作用メカニズム
2.東京医科歯科大学時代
Wisconsin 大学から日本に戻って、私は活性型ビタミンDの臨床応用のための基礎研究に着手した。そのきっかけを与えてくれたのは東京慈恵会医科大学上田内科の小椋陽介先生と川口良人先生(腎臓内科学)であった。当時、日本には2万人を超える慢性腎不全患者さんが居て、人工血液透析療法が普及し始めた頃であった。腎不全を患う患者さんの多くは貧血と骨の病気に悩んでいた。これは、腎臓における赤血球を作るエリスロポエチンと骨の代謝に関係する活性型ビタミンDの産生不足に由来すると考えられた。私は、腎不全に由来する骨の病気(腎性骨異栄養症)を患う患者さんに活性型ビタミンDを投与すれば骨の症状が改善するのではないかと考え、東京医科歯科大学医用機材研究所の金子主税先生(後に東北大学薬学部教授)と共に活性型ビタミンDの有機合成に挑戦した。しかし、当時の技術では化学的にビタミンDの25位に水酸基を導入する良い化合物を見つけることができず、やむなく1α位だけに水酸基を導入したビタミンD誘導体 [1α(OH)D3] を有機合成した。両側の腎臓を摘出したラットに1α(OH)D3を投与したところ、血清Ca濃度が顕著に上昇することを見出した。この知見によって、1α(OH)D3は1α,25(OH)2D3の代替物質となり得ると確信し、中外製薬と共同して新薬の開発にチャレンジすることになった。この時、中外製薬側の1α(OH)D3開発のプロジェクトリーダ―に選ばれたのが西井易穂さんであった。西井さんは大変優れた生化学者で、日本の生化学の世界では顔を良く知られる有名人であった。西井さんと共に、1α(OH)D3が肝臓で速やかに1α,25(OH)2D3に変換することを証明した肝還流実験(1975年)[図6]と、柴田腎炎の病態モデルラットを作製して1α(OH)D3が腎炎に基づく骨の疾病を改善することを明らかにした動物実験[1978年]は記憶に新しい。1α(OH)D3 の25位を水酸化する酵素は、奥田九一郎先生(広島大学歯学部)がすでに胆汁酸の代謝研究で発見していたミトコンドリアの酵素CYP27A1であった。ビタミンD2(3)の生理的な25-水酸化酵素はミクロソームの酵素(CYP2R1)で、この二つの酵素タンパクの25-水酸化反応を比べると、CYP27A1による1α(OH)D3の25-水酸化反応は基質濃度に依存せず直線的に進行するのに対し、CYP2R1によるビタミンD3の25-水酸化反応は基質濃度依存的に調節されていることが判明した[図6、挿入図]。こうして出来上がった新薬がAlfacalcidol (商品名:アルファロール)で、アルファロールはその後7年間の開発期間を経て1981年、腎性骨異栄養症、二次性副甲状腺機能亢進症、ビタミンD抵抗性クル病などに適用が認可され、1983年には骨粗鬆症にも適用が拡大された[図7]。Alfacalcidol はBisphosphonate製剤が出現するまで骨粗鬆症治療薬の主力の薬剤として広く使用され、毎日100万人の患者さんが服用して下さる時期もあった。現在では骨粗鬆症治療薬の主力はBisphosphonate 製剤や新しいビタミンD製剤 (Eldecalcitol)などによって取って代わられている[図8]。
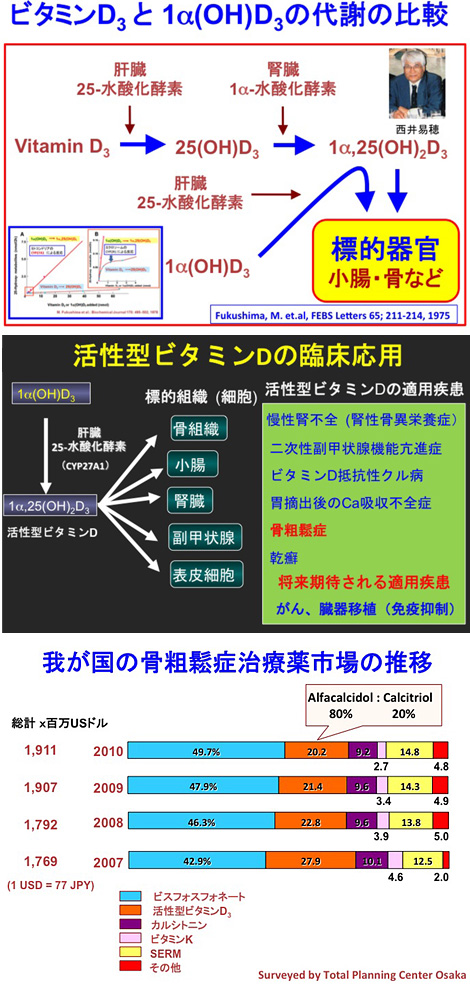
(上:図6)ビタミンD3と1α(OH)D3 の25-水酸化反応の比較 (1975, 1978)
(中:図7)活性型ビタミンDの臨床応用
(下:図8)我が国の骨粗鬆症治療薬市場の推移
私が産学連携研究の重要性を認識したのはWisconsin大学留学中に知ったWARF (Wisconsin Alumni Research Foundation) の存在であった。DeLuca先生の前任者であったSteenbock教授は、1924年、食品に紫外線を照射すると骨の病気[クル病]を予防・治癒する効果が生まれることを発見した。この発見に基づいて、米国食品医薬局 (FDA) は米国で市販される牛乳にはすべてビタミンDを添加しなければならないという取り決めを行った(Vitamin D milk)。その結果、Wisconsin大学には莫大な特許料が入ることになったが、Steenbock先生はそれを全額出費してWisconsin大学の基礎研究を支援するための財団WARFを設立し、大学で生み出される特許を一括管理するシステムを確立した。米国では、100年近くも前にこのような財団が創設され、研究成果を社会へ還元する活動をしていることを知った。日本で産学連携研究の重要性が注目されるようになったのはこの10-15年のことで、私達がAlfacalcidol の創薬研究を行っていた頃は産学連携研究に対する雰囲気は必ずしも温かいものではなかった。
3.昭和大学時代
1977年、昭和大学歯学部の開設に伴い、歯学部生化学教室の初代教授として着任した私は、堀内 登、阿部 悦子、高橋 直之、宮浦 千里, 宇田川 信之らと共に「骨とビタミンD」の研究を始めることになった。まず、1981年、阿部悦子(後にNY. Mount Sinai Medical Center 研究教授)は、活性型ビタミンDがマウスやヒトの骨髄性白血病細胞(M1, HL-60) の増殖を抑制し、マクロファージへの分化を選択的に誘導する強力な作用を持つことを報告した[図9]。

(図9)ビタミンDの細胞分化誘導作用の発見 (PNAS,USA, 1981)
この論文(PNAS, USA 78, 4990-4994,1981)はビタミンDの生理作用をCa ・骨内分泌学の世界からbeyond boneの領域へと広げるきっかけを与えた。またこの論文は、今まで、ビタミンD誘導体の生理活性の測定が専らin vivoのアッセイ系に限られていて、一週間以上の時間を要していたのに対し、ビタミンDの細胞分化誘導作用の発見によって、僅か2-3日間の短時間でビタミンD誘導体の活性測定をin vitroで可能にした。その結果、22-oxa-1α,25(OH)2D3 (Oxarol), 1α,25R(OH)2D3 (Tacalcitol)、Betametasone dipropionate (Calcipotriol)、2β-(3-hydroxyoropoxy)-1α,25(OH)2D3(Eldecalcitol) などの新規ビタミンD誘導体が続々と登場し、乾癬、副甲状腺機能亢進症、骨粗鬆症などの治療薬として適用されるようになった。続いて、1983年、阿部は1α,25(OH)2D3が直接、あるいはリンパ球を介して肺胞マクロファージの融合を促進し、多核巨細胞を形成させる作用を持つことを報告した (PNAS, USA 80, 5583-5587, 1983)。この論文がきっかけになって、私どもの興味は「破骨細胞形成とビタミンD」の研究へと向かうことになった。
破骨細胞は骨組織にのみ存在し。石灰化した骨組織を破壊・吸収する唯一の細胞である。破骨細胞は骨髄あるいは血液中に存在するマクロファージ系の造血細胞から形成されるが、マクロファージは身体中至る所に存在するのに、なぜ破骨細胞は骨組織にしか存在しないのだろうか。直橋直之と宇田川信之(いずれも後に松本歯科大学教授)はその鍵を握っているのは骨組織にのみ存在する骨芽細胞ではないかと考えた。この仮説の根拠になったのはGideon Rodan先生 とJack Martin先生がCalcified Tissue Int. (33; 349-351, 1981) に報告した一つの論文であった。 Rodan先生とMartin 先生の論点は、[1]PGE2, PTH, ビタミンDなどin vivo で骨吸収活性を示すホルモンやサイトカインの受容体が破骨細胞にはなく、骨芽細胞に存在すること、[2]これらの骨吸収因子の骨芽細胞における受容体への親和性がin vivoで骨吸収を誘導する活性と密接に相関することの2点であった[図10]。
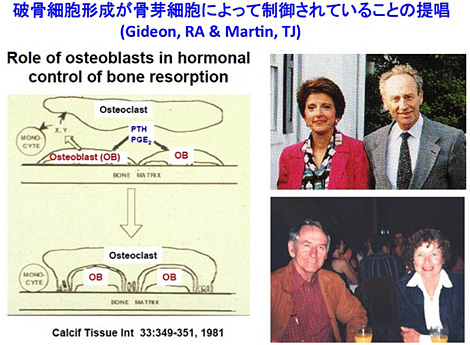
(図10)破骨細胞形成が骨芽細胞によって制御されていることを最初に示唆したRodan先生とMartin先生の論文(Calcified Tissue Int., 1981)
高橋と宇田川は破骨細胞形成における骨芽細胞の役割を明らかにするために、骨芽細胞と脾細胞(破骨細胞前駆細胞)を1α,25(OH)2D3の存在下で共存培養するin vitro の実験系を構築した(Endocrinology 123, 2600-2602, 1988)。この共存培養系で、破骨細胞は骨芽細胞と脾細胞が共存する場合のみ1α,25(OH)2D3に依存して形成されることが判明した[図11]。この共存培養系では、in vivo で骨吸収活性を持つPTH , PGE2, IL-1, IL-6 にも反応して破骨細胞が形成された。また、破骨細胞形成には骨芽細胞と脾細胞のcell--cell contact が必要であった。この共存培養系による破骨細胞形成法は、その後、再現性の高いin vitroの破骨細胞形成系として, 世界の標準的な実験法となった。我々はこの共存培養系の実験成績を基にして、1992年、骨吸収活性を持つ様々なホルモンやサイトカインが破骨細胞を形成させるためには、まずそれらは骨芽細胞を標的として、その細胞膜上に膜結合型の破骨細胞分化因子(Osteoclast differentiation factor, ODF) を発現させるという作業仮説を提唱した(Endocrine Reviews 13; 66-80, 1992)。破骨細胞形成を促進する因子はシグナル伝達の上から、[1]VDRを介するもの、[2]cAMPを介するもの、[3]gp130を介するものの3種類に分けられた。それらのシグナルは全て骨芽細胞上に膜結合型サイトカインODFを誘導すると考えられた。
1997年、雪印乳業生物科学研究所の東尾侃二博士が率いる研究グループは、共存培養系で破骨細胞形成を強く抑制する新しいタンパク質の分子クローニングに成功した。この因子はヒト胎児線維芽細胞 (IMR90) の培養上清から単離され、破骨細胞形成抑制因子(Osteoclastogenesis inhibitory factor, OCIF) と名付けられた。 OCIFはcysteine-rich domainを4個持つ新規の分泌性TNF受容体であった。丁度同じころ、米国のAmgen 社も新規TNF受容体をクローニングし、そのTransgenic mouse を作成した。このマウスは破骨細胞が全く形成されない典型的な大理石骨病の症状を呈することが判明した。Amgen社はこの因子には骨を守る作用があることから、Osteoprotegerin (OPG) と命名した。OPGとOCIFは同一のタンパク質であった。
OCIF/OPGが骨芽細胞を標的として、ODF による破骨細胞形成を強力に抑制するという実験成績に基づき、雪印乳業の東尾グループと我々は1α,25- (OH)2D3で処理した骨髄性間質細胞(ST2)のcDNAライブラリーからOCIFと特異的に結合するタンパク質の分子クローニングを行った。その結果、この分子こそ我々が長い間追い求めてきた骨芽細胞の産生する破骨細胞分化因子(ODF)そのものであった。ODFは316個のアミノ酸からなる膜貫通領域を持つTNFリガンドファミリーに属する新規タンパク質であった(PNAS,USA 95; 3597-3602)[図12]。
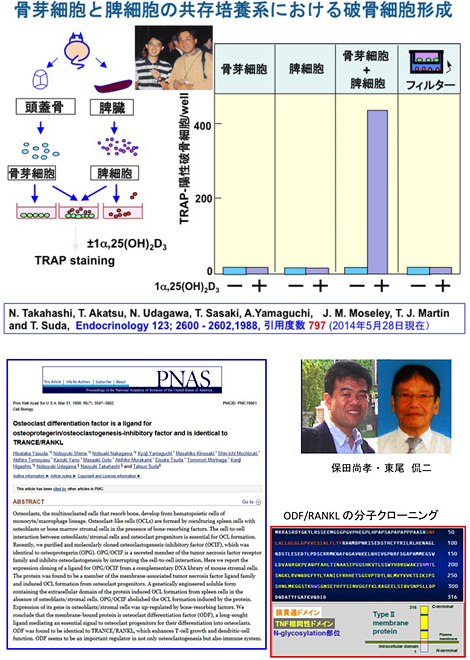
(上:図11)骨芽細胞と脾細胞の共存培養系で1α,25(OH)2D3に依存して破骨細胞を作った最初の論文(Endocrinology, 1988)
(下:図12)ODF/RANKLの分子クローニングを報じたPNAS, USA論文(1998)
また、ODFの膜結合ドメインのC末端側で切断した可溶性ODF(sODF)とM-CSF を添加することによって、骨芽細胞がなくても脾細胞や骨髄マクロファ―ジから破骨細胞が形成されること、形成された破骨細胞は実際に骨や象牙質を破壊・吸収する活性を持つことが確かめられた。このPNAS論文は1998 年のRed Hot Paper の第32位にランクされ、これまでの引用度数は3,200 を超えた。ODFは、骨の研究領域ではAmgenがOPGの結合タンパクとしてクローニングしたOPGL、 また、免疫学の領域の2つの研究グループから発見されたRANKL (Receptor activator of NF-kB ligand)とTRANCE[TNF-related activation-induced cytokine]と同一物質であった。米国骨代謝学会(ASBMR) は、今後、破骨細胞形成系のLigand はRANKL, 受容体はRANK, RANKLとRANKの結合を阻止するおとりの受容体はOPGという名前を使用した論文だけを受理することを決めた。
破骨細胞形成系(RANKL/RANK/OPG)の発見によって、破骨細胞の形成と機能を調節する骨芽細胞の役割は分子レベルで理解されるようになった。すなわち、破骨細胞形成における骨芽細胞の役割はM-CSFとRANKLを産生することである。M-CSFは骨芽細胞から骨吸収因子を必要とせず自律的に産生されるが、RANKLはPTH, PGE2, IL-1, IL-6, 1α,25(OH)2D3などの骨吸収因子の刺激によって骨芽細胞から産生される。M-CSFが破骨細胞前駆細胞のM-CSF受容体 (c-Fms) に結合すると、その細胞膜上にRANKL受容体(RANK)が誘導される。RANKLは破骨細胞前駆細胞に誘導されたRANKに結合して前駆細胞を破骨細胞に分化させると共に、それを活性化させ、成熟破骨細胞を作る。OPGはRANKLとRANKの結合を切断するおとりの受容体(decoy receptor) である[図13]。OPGは最初、骨吸収の抑制する本命の治療薬として創薬することが期待されたが、血液中の半減期が短いため、治療薬にすることはできなかった。 代わりに、AmgenはRANKL 抗体(Denosumab) を創製した。Denosumabは骨粗鬆症や乳がんの骨転移におけるRANKL誘導性の骨吸収を特異的に抑制する強力な抗体医薬となった。
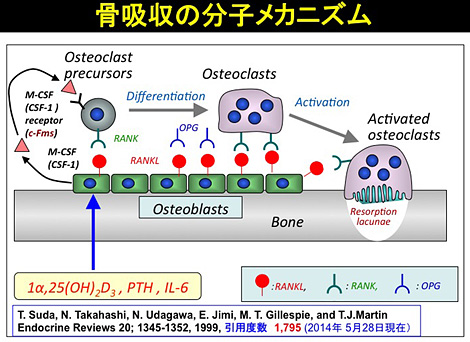
(図13)解明された破骨細胞形成と機能発現の分子メカニズム(Endocrine Reviews, 1999)
おわりに
ビタミンDは、一世紀も前に骨の形成と石灰化を促進する骨にとって良いビタミン(anti-rachitic agent)として発見された。しかし、100年後の現在も、ビタミンDが骨芽細胞による骨形成と石灰化を直接促進することを示す実験的証拠は得られていない。 むしろ逆に、in vitroでは、活性型ビタミンDは骨芽細胞に働いて破骨細胞誘導因子(RANKL)の産生を促進し、骨吸収を促進する強力な作用を持つことが明らかにされている。つまり、ビタミンDは骨にとって悪いビタミンのように見える。このin vivo とin vitro の矛盾はどのように説明すればよいであろうか。一つの説明は、骨芽細胞のRANKL産生と小腸でのCa輸送に及ぼす1α,25(OH)2D3の所要量に関係しているように思われる。In vitro でRANKLを産生させるのに要する1α,25(OH)2D3の濃度は健常人の1α,25(OH)2D3の血清濃度(10-10 M)の10倍(10-9 M) から100倍(10-8 M)の高濃度が必要である。生理量の活性型ビタミンDの投与ではin vivoではむしろ骨吸収は抑制されることが知られている。これは、生理濃度の活性型ビタミンDではまず小腸 (十二指腸) でのCa輸送が亢進し、血清Ca濃度を上昇させ、間接的に骨の石灰化を促進していると思われる。1α,25(OH)2D3による血清Caの上昇はPTHの分泌抑制を齎すことも知られている。ビタミンDの大量投与時やビタミンD中毒の時に起こる高Ca血症の存在は、ビタミンDが用量によって破骨細胞形成に関与している証拠である[図14]。
最初の生命体は海水中で誕生し、動物は河口→淡水→陸上へと棲息場所を移して行った。この棲息環境の推移に伴って最も激しく変動したのは棲息環境のCa濃度と重力であった。海水のCa濃度(10 mM)は高等脊椎動物の血清のCa 濃度 (2.5 mM) の4倍あり、血清のこのCaホメオスタシスを維持するために脊椎動物は骨組織をCaの貯蔵庫として利用するようになった。小腸からのCaの供給が断たれても脊椎動物が血清Caの恒常性を維持できるのは骨組織からのCa の供給[骨吸収]があるためである。陸棲脊椎動物にとって骨組織は海そのものである。血清Caの恒常性維持のために中心的な役割を果たすのはビタミンDである。そのために、in vivo で骨を良くする栄養素として誕生したビタミンDが陸棲脊椎動物では破骨細胞分化(RANKL)を誘導する役割を持つようになったと思われる。椎動物が生きてゆくために、生体のCaホメオスタシスを維持することは骨を犠牲にしても果たさなければならない必須の仕事であった。In vivoではビタミンDの投与によって骨吸収が抑制され、骨形成(石灰化)が促進されるが、その分子レベルのメカニズムの解明は今後の研究に待つところが大きい[図15]。
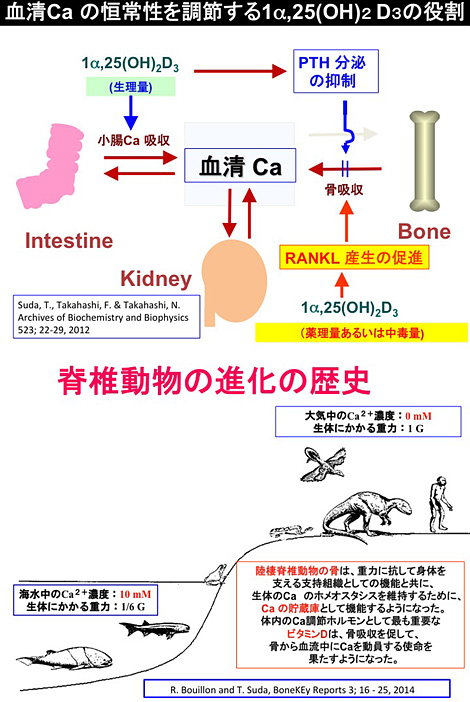
(上:図14)血清Caの恒常性を調節する1α,25(OH)2D3 の役割 (ABB, 2012)
(下:図15)脊椎動物の進化の歴史と骨の役割 (BoneKEy Reports, 2014)
終わりに、これまで半世紀以上に亘り私が基礎研究を続けることができたのはひとえに先輩諸氏のご指導、多くの共同研究者の皆さんのご努力、それに中外製薬を初めとする製薬企業の皆様の産学連携のお蔭である。この場を借りて、永年に亘る皆様のご指導と熱き友情に心から御礼を申し上げる。
次世代への一言
私はDeLuca 先生からビタミンD代謝のサイエンスを学ぶと共に、「Science is fun」という言葉を教えていただいた。科学することはなぜそれほど楽しいのか。私の解釈は、
[1]科学はまだ誰も知らない新知見を発見する喜びを与えてくれる、
[2]科学は研究者を平等に扱ってくれる、
[3]科学は世界中に友達を作ってくれる、
[4]科学は自分の研究が社会に役立つ喜びを与えてくれる、
などなどです。
この言葉を若い研究者の皆さんに贈ります。