





TOP > 1st Author > 大津 圭史
Semaphorin4D-RhoA-Akt経路はエナメル芽細胞分化における細胞極性、エナメルタンパク発現を統合的に制御する
The Semaphorin 4D-RhoA-Akt signal cascade regulates enamel matrix secretion in coordination with cell polarization during ameloblast differentiation
| 著者: | Keishi Otsu, Hiroko Ida-Yonemochi, Naoki Fujiwara, Hidemitsu Harada |
|---|---|
| 雑誌: | Journal of Bone and Mineral Research, 2016 Nov;31(11):1943-1954. doi: 10.1002/jbmr.2876. |
- エナメル芽細胞
- Semaphorin4D
- Rhoシグナリング

右:著者、左:原田英光教授
論文サマリー
歯の表面を覆うエナメル質は体の中でもっとも硬い組織であり、食事の摂取や咀嚼はもちろん、外部刺激から歯髄を保護する役割を担っている。このエナメル質形成は,エナメル芽細胞とよばれる外胚葉系上皮細胞が担っており,骨芽細胞のような間葉系細胞の石灰化とはまったく異なるメカニズムによって行われる。上皮細胞は,細胞間の連携を保ちながら,増殖による細胞数の増加と基質の分泌や石灰化のための形態変化を行う。このような上皮細胞独特な分化過程を制御する分子メカニズムは十分に理解できていない。我々はRhoシグナルを中心にエナメル芽細胞の分化メカニズムを長年にわたって研究しており,今回は上流と下流のシグナルカスケードの探索と機能解析によって上皮細胞の分化過程でのSemaphorin(Sema)シグナルの役割を新たに発見した。
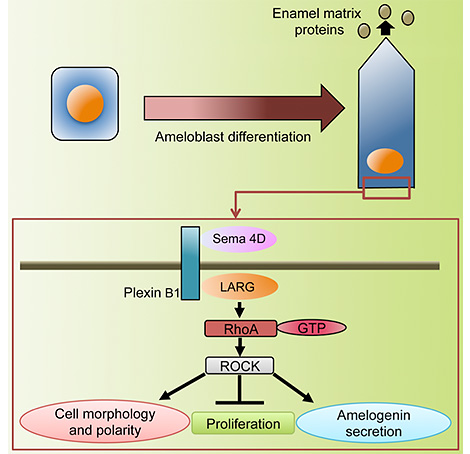
Sema4Dとその受容体PlexinB1、活性化型RhoAは同様の発現局在を示しながら,エナメル芽細胞の分化するに伴って増強する。培養細胞や器官培養を用いた実験などから、Sema4D-PlexinB1シグナルがRhoAを活性化し、アクチン重合、細胞接着因子やエナメルタンパク(アメロゲニン)の発現を促進すること,さらにSema4D-PlexinB1によるRhoAの活性化は、グアニンヌクレオチド交換因子LARGを介して起こっていること, RhoAの活性化の下流でAktをリン酸化して細胞増殖やアメロゲニンの分泌を制御している事がわかった。エナメル上皮細胞特異的にRhoAのドミナントネガティブフォームが発現する,RhoAの機能喪失遺伝子改変マウスでも、エナメル芽細胞の細胞形態、極性の変化が起こらず、アクチン重合、細胞接着因子の低下、アメロゲニンの発現抑制と分泌方向の異常が見られ,生体でも同様のメカニズムが存在することを裏付けた。以上の結果から、Sema4D-RhoA-Akt経路が、細胞形態・極性変化、細胞増殖、エナメルタンパク分泌を統合的に制御する新規の上皮分化メカニズム(下図)が明らかとなった。
著者コメント
現所属の岩手医科大学に赴任してから、歯の発生と再生研究に取り組んでいます。今回の研究ではエナメル芽細胞の分化を制御するメカニズムに焦点をあて、その一端を明らかにすることができました。今回の結果は、エナメル質形成不全の原因の究明、発症を予防する薬の開発に役立つとともに、幹細胞を用いたエナメル質再生に応用できると期待されます。
歯・歯周組織の発生では、細胞の分化、増殖、移動などが時空間的に精密にコントロールされ、複数の硬組織(エナメル質、象牙質、セメント質、歯槽骨)と軟組織(歯髄、歯根膜)を合わせ持つ複雑な構造が作り上げられていきます。これらを統合的に理解するには、まだまだ明らかにしなければならない課題が多く残されていますので、今後も地道な研究を積み重ねていきたいと考えています。
最後になりましたが、本研究を発表するにあたり、私の遅々として進まない研究を辛抱強く見守り、指導してくださった原田英光教授をはじめ、多大なご協力を賜りました共著者の先生方、教室員の皆様にこの場を借りて深く感謝申し上げます。そしていつも私の仕事を応援し、支えてくれる妻と子供達に心から感謝します。(岩手医科大学 解剖学講座 発生生物・再生医学分野・大津 圭史)